
Get your Free Calisthenics Certification
100% Online Training — Study Wherever You Are
What You'll Get
Discover what makes our approach unique
Online quizzes
Online quizzes to practice your knowledge
Flexible learning
Live or self-paced learning
Open to everyone
No prior knowledge or degree required
Popular Articles
Our most popular articles to get you started

How to Fix Tight Hip Flexors: Best Stretches, Exercises & Daily Routine
Complete guide to relieving tight hip flexors with evidence-based stretches, strengthening exercises, and a daily routine. Learn causes, symptoms, self-tests, and how tight hips affect your calisthenics training.

Calisthenics for Beginners: Complete 30-Day Program
Start your calisthenics journey with this free 30-day beginner workout plan. Build strength, improve mobility, and master fundamental bodyweight exercises with step-by-step progressions.

How to Fix Anterior Pelvic Tilt: Evidence-Based Exercises, Stretches & Posture Correction
Complete guide to correcting anterior pelvic tilt with proven exercises, stretches, and lifestyle modifications. Learn the anatomy, causes, and step-by-step rehabilitation protocol.
Featured Courses
Professional certification programs to advance your career
Popular Lessons
Start learning with our most popular content
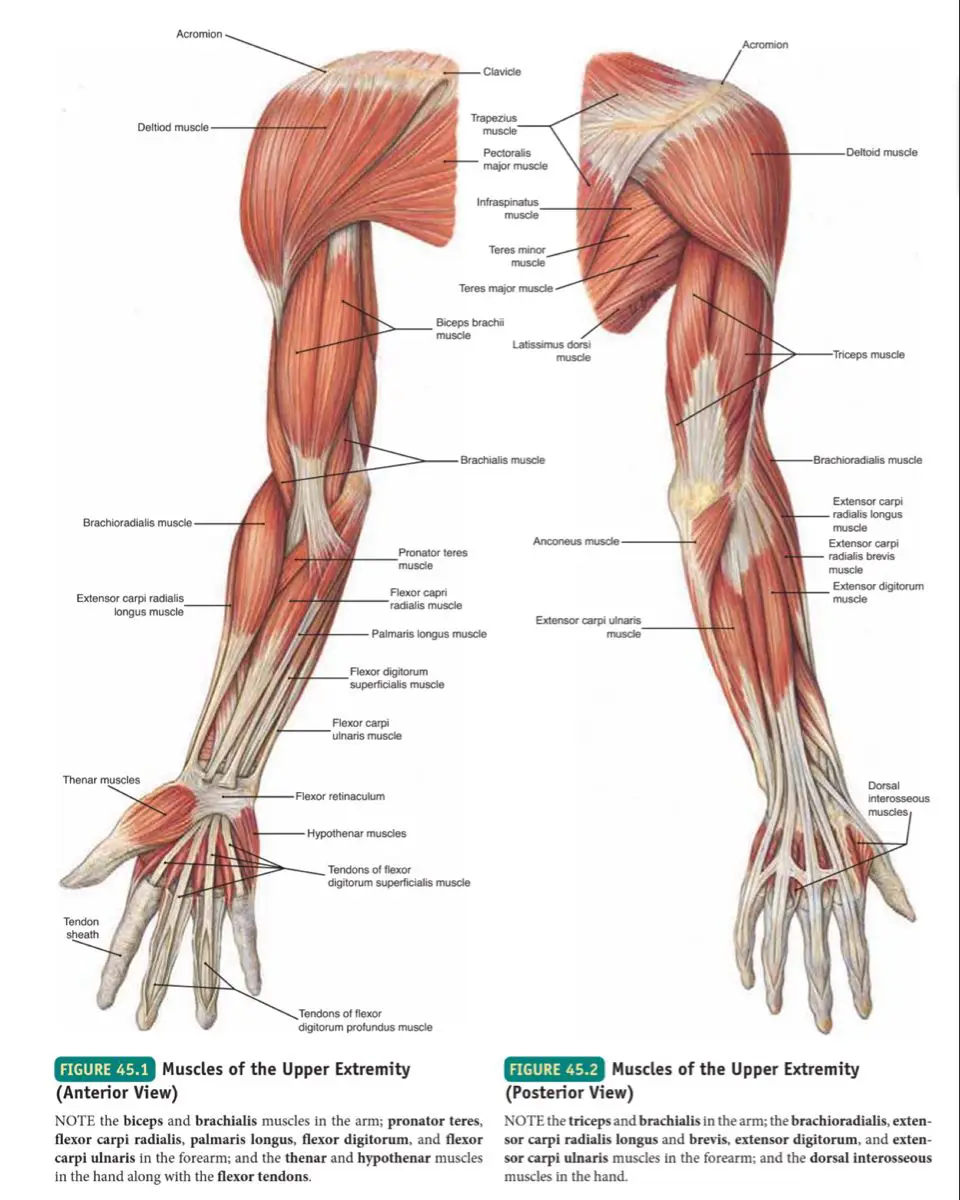
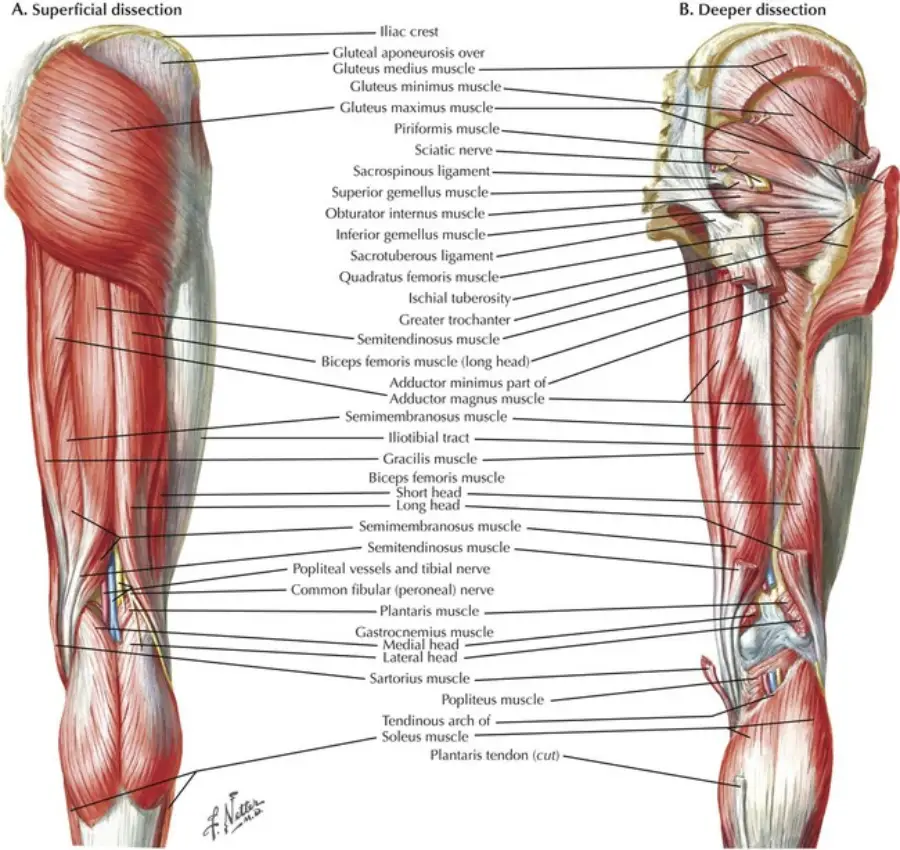
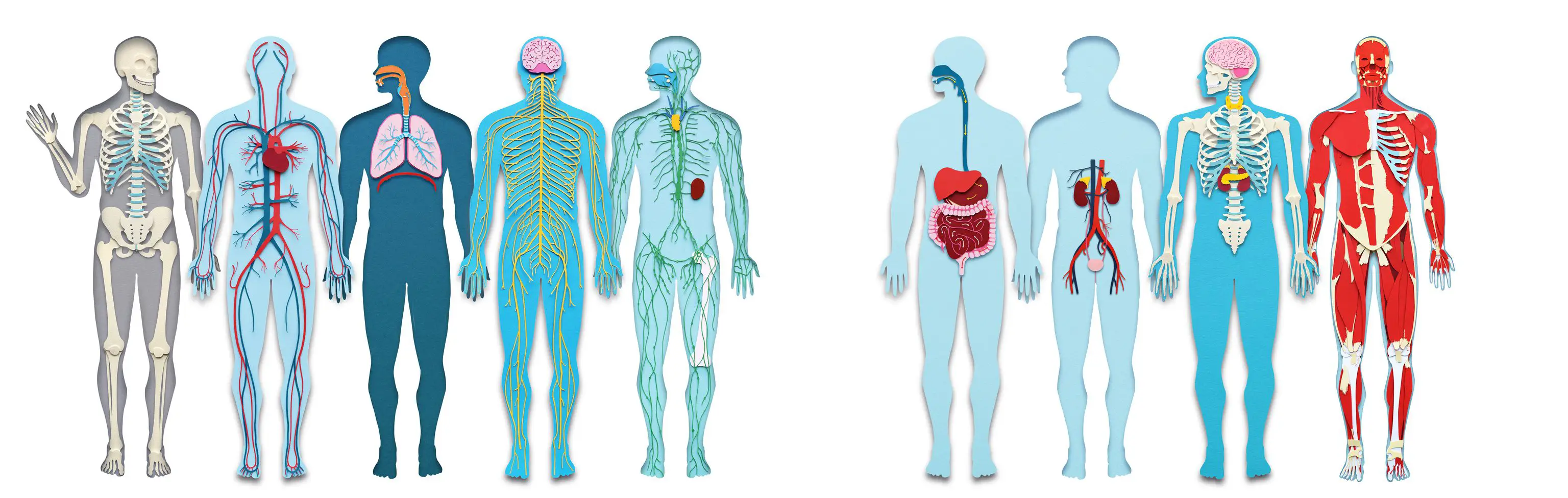
What You'll Learn
Complete curriculum covering everything from anatomy to advanced programming
Anatomy & Kinesiology
- Musculoskeletal system fundamentals
- Biomechanics and movement patterns
- Injury prevention and rehabilitation
Training Methodology
- Progressive overload principles
- Program design and periodization
- Exercise progressions and regressions
Exercise Science
- Muscle physiology and adaptation
- Energy systems and metabolism
- Recovery and nutrition basics
Professional Skills
- Client assessment techniques
- Coaching and communication
- Safety and risk management
Latest Articles
Evidence-based fitness insights

30-Minute Full-Body Calisthenics Workout (Do It Today)
A complete 30-minute full-body calisthenics workout you can do today. Warm-up, a 20-minute push, pull, legs, and core circuit, plus a cooldown, with a regression and progression for every move.
Read More
The 4-Week Summer Body Calisthenics Plan (No Gym, No Equipment)
A complete 4-week summer body calisthenics plan with weekly schedules, exact sets and reps, and beginner-to-intermediate progressions. Build a lean, athletic physique using only your bodyweight.
Read More
AI Tools Every Calisthenics Coach Should Use in 2026
A practical 2026 guide to the AI tools that save calisthenics coaches hours every week — workout programming, client check-ins, social media, onboarding automation, and nutrition planning. With recommended prompts and a free AI course for fitness coaches.
Read MoreFrequently Asked Questions
About Calisthenics Association
Unlock your potential with the first online school offering FREE Calisthenics training certification! Our comprehensive, fully online courses allow you to become a certified Calisthenics trainer from anywhere in the world. Join thousands of students across the globe who have transformed their lives and careers with our expert training. Why wait? Start your journey with us today and embrace the future of fitness training. The world of Calisthenics awaits you!