Κεφάλαιο 6 – Φυσιολογία των μυών
Δομή των σκελετικών μυών
Οι σκελετικοί μύες αποτελούν το 40% περίπου του σωματικού βάρους στους άνδρες και το 32% στις γυναίκες, με τους λείους μυς και τον καρδιακό μυ να αποτελούν το υπόλοιπο 10%. Μέσω της συστολής των μυών αναπτύσσεται τάση, επιτελούνται οι κινήσεις του σώματος και παράγεται έργο.
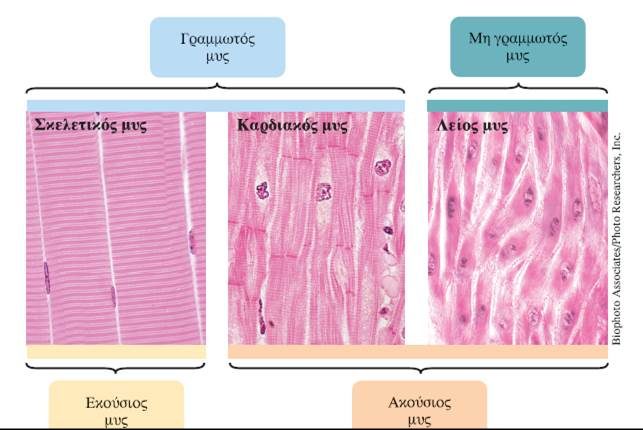
Οι τρεις τύποι του μυϊκού ιστού ταξινομούνται με δύο τρόπους σύμφωνα με τα κοινά χαρακτηριστικά τους. (1) Οι σκελετικοί μύες και ο καρδιακός μυς είναι γραμμωτοί, ενώ οι λείοι μύες είναι μη γραμμωτοί. (2) Οι σκελετικοί μύες είναι εκούσιοι, δηλαδή ελέγχονται από τη θέλησή μας, ενώ ο καρδιακός μυς και οι λείοι μύες λειτουργούν ακούσια, δηλαδή ανεξάρτητα από τη θέλησή μας.
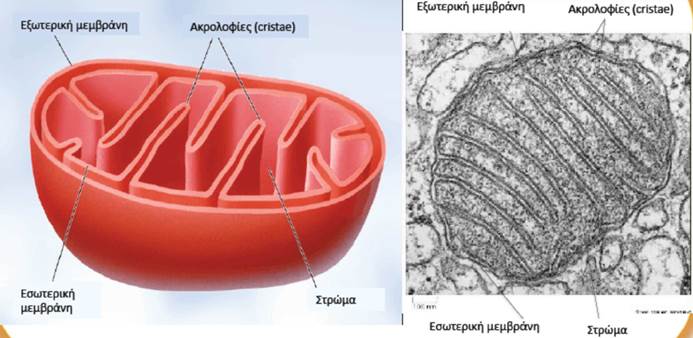
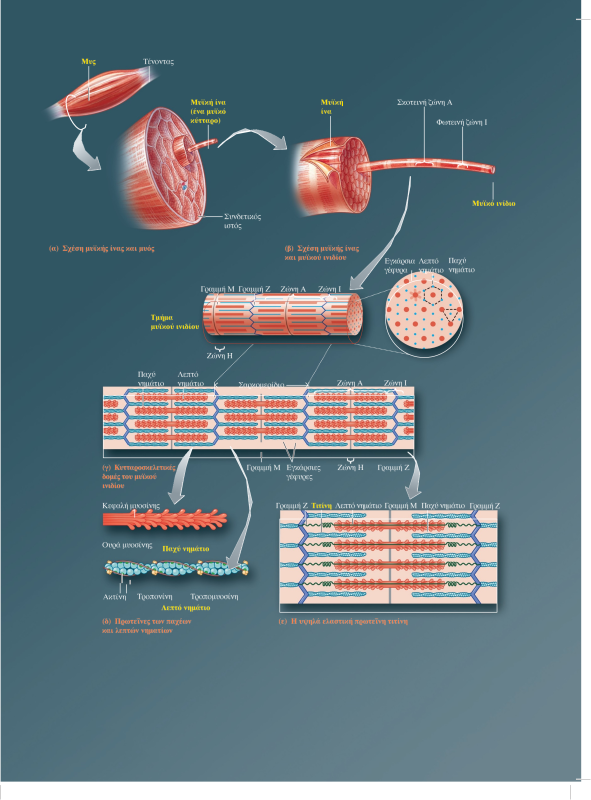
Οι σκελετικοί μύες αποτελούνται από δέσμες επιμήκων, κυλινδρικών μυϊκών κυττάρων (που ονομάζονται μυϊκές ίνες), οι οποίες περιβάλλονται από συνδετικό ιστό. Κατά τη διάρκεια της εμβρυϊκής ανάπτυξης, οι σκελετικές μυϊκές ίνες σχηματίζονται από την συγχώνευση πολύ μικρότερων κυττάρων που ονομάζονται μυοβλάστες. Κάθε μυϊκή ίνα διαθέτει πολλούς πυρήνες και μεγάλο αριθμό μιτοχονδρίων, καθώς τα μιτοχόνδρια είναι τα κυτταρικά οργανίδια στα οποία παράγεται η ενέργεια που απαιτείται για την έντονη δραστηριότητα των σκελετικών μυών.
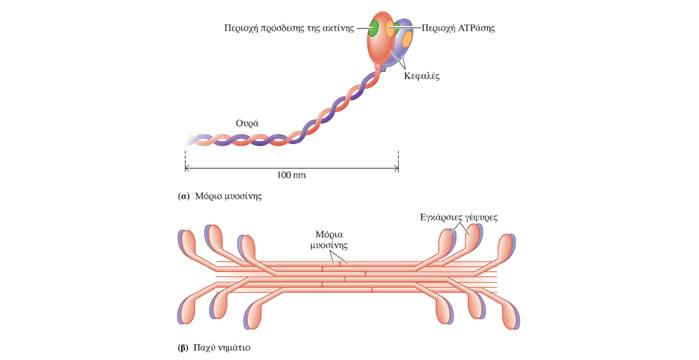
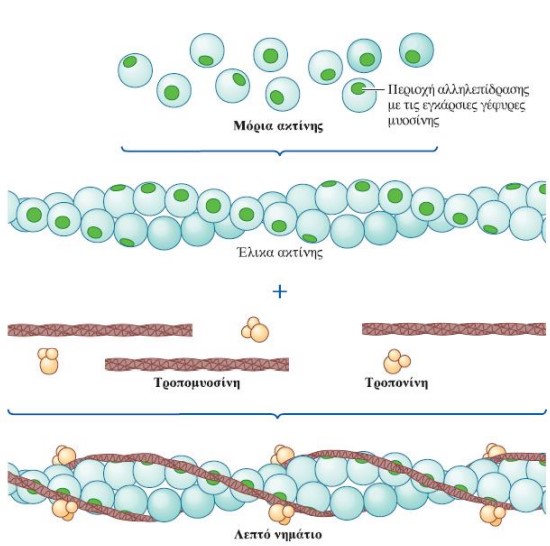
Κάθε μυϊκή ίνα περιέχει μεγάλο αριθμό μυϊκών ινιδίων (εξειδικευμένες συσταλτές δομές οι οποίες αποτελούν το 80% του συνολικού όγκου της μυϊκής ίνας), καθένα από τα οποία αποτελείται από εναλλασσόμενες σειρές μερικώς επικαλυπτόμενων παχέων και λεπτών νηματίων. Η διάταξη των παχέων και των λεπτών νηματίων έχει ως αποτέλεσμα την εναλλαγή σκοτεινών ζωνών Α και φωτεινών ζωνών Ι κατά μήκος των μυϊκών ινιδίων. Στην εναλλαγή αυτή οφείλεται η γραμμωτή εμφάνιση των σκελετικών μυϊκών ινών στο μικροσκόπιο. Η περιοχή ανάμεσα σε δύο γραμμές Ζ αποτελεί την λειτουργική μονάδα του σκελετικού μυός και ονομάζεται σαρκομερίδιο. Τα παχέα νημάτια αποτελούνται από την πρωτεΐνη μυοσίνη. Οι εγκάρσιες γέφυρες σχηματίζονται από τις σφαιρικές κεφαλές των μορίων μυοσίνης, οι οποίες προβάλλουν από κάθε παχύ νημάτιο προς τα λεπτά νημάτια που το περιβάλλουν. Τα λεπτά νημάτια αποτελούνται κυρίως από την πρωτεΐνη ακτίνη, η οποία φέρει θέσεις που αλληλεπιδρούν με τις εγκάρσιες γέφυρες της μυοσίνης προκαλώντας μυϊκή συστολή. Σε κατάσταση ηρεμίας δύο άλλες πρωτεΐνες που βρίσκονται στην επιφάνεια του λεπτού νηματίου, η τροπονίνη και η τροπομυοσίνη, παρεμποδίζουν την αλληλεπίδραση της ακτίνης με τις εγκάρσιες γέφυρες της μυοσίνης. Η τροπομυοσίνη και η τροπονίνη ονομάζονται συχνά ρυθμιστικές πρωτεΐνες (regulatory proteins) λόγω του ρόλου τους στην κάλυψη (παρεμποδίζοντας τη μυϊκή συστολή) ή στην αποκάλυψη (επιτρέποντας τη μυϊκή συστολή) των θέσεων αλληλεπίδρασης ακτίνης-μυοσίνης. Μεμονωμένες αλυσίδες μιας γιγαντιαίας, ιδιαίτερα ελαστικής πρωτεΐνης, η οποία ονομάζεται τιτίνη (titin), ξεκινώντας από τη γραμμή M εκτείνονται και στις δύο κατευθύνσεις κατά μήκος των παχέων νηματίων, προς τα αντίθετα άκρα του σαρκομεριδίου, δηλαδή προς τις δύο γραμμές Z που το οριοθετούν. Η τιτίνη, που αποτελείται από περίπου 30.000 αμινοξέα, είναι η μεγαλύτερη πρωτεΐνη του ανθρώπινου οργανισμού και μαζί με τις πρωτεΐνες της γραμμής Μ, βοηθά στην σταθεροποίηση της θέσης των παχέων νηματίων σε σχέση με τα λεπτά νημάτια -συμβάλλοντας στην δομική σταθερότητα του σαρκομεριδίου-, λειτουργώντας σαν ελατήριο, αυξάνει κατά πολύ την ελαστικότητα των μυών (παράλληλο ελαστικό στοιχείο (parallel-elastic component) των μυών) και συμμετέχει στη μετάδοση σημάτων.
Μυς (όργανο) → μυϊκή ίνα (κύτταρο) → μυϊκό ινίδιο (εξειδικευμένη ενδοκυτταρική δομή) → παχέα και λεπτά νημάτια (στοιχεία του κυτταροσκελετού) → μυοσίνη και ακτίνη (πρωτεΐνες)

![]()
Μοριακή βάση της συστολής των σκελετικών μυών
Η διέγερση της σκελετικής μυϊκής ίνας από τον κινητικό νευρώνα επάγει τη συστολή της. Κατά τη συστολή, διαδοχικοί κύκλοι πρόσδεσης στην ακτίνη και κάμψης των εγκάρσιων γεφυρών έλκουν τα λεπτά νημάτια στο εσωτερικό του σαρκομεριδίου. Η αλληλεπίδραση των εγκάρσιων γεφυρών μυοσίνης με την ακτίνη προκαλεί τη συστολή των μυών μέσω ενός μηχανισμού διολίσθησης των νηματίων. Κάθε εγκάρσια γέφυρα μυοσίνης φέρει δύο διακριτές λειτουργικές περιοχές: μία θέση πρόσδεσης της ακτίνης και μία θέση με δραστικότητα ATPάσης.
[Αντλία νατρίου-καλίου ή ATPάση Na-K:
Η αντλία νατρίου-καλίου ή ATPάση Na-K είναι χαρακτηριστικός μηχανισμός ενεργητικής μεταφοράς. Πρόκειται για διαμεμβρανική πρωτεΐνη, η οποία για κάθε 3 ιόντα Να+ που εξάγει από το κύτταρο εισάγει 2 ιόντα Κ+. Η άντληση αυτή γίνεται με την υδρόλυση ATP σε ADP. Η αντλία δουλεύει κυκλικά και είναι κανάλι διπλής κατεύθυνσης, αλλά επιτρέπει την έξοδο μόνο σε ιόντα Νa και την είσοδο μόνο σε ιόντα Κ. Την ενέργεια για τη λειτουργία αυτή, την εξασφαλίζει από την υδρόλυση της ATP (τριφωσφορικής αδενοσίνης ή τριφωσφαταδενοσίνης):
ATP + H2O → ADP + H3PO4 + ΔG
Έτσι προκύπτει η ADP, ενώ απελευθερώνεται ενέργεια: ΔG = 30,5kJ/mol
[mol ή mole = γραμμομόριο, μονάδα μέτρησης για την ποσότητα της ύλης ενός σώματος] [Η μεταβολή της ελεύθερης ενέργειας Gibbs (ΔG) είναι το άμεσο κριτήριο για την αυθόρμητη επιτέλεση ενός πειράματος (χημικής αντίδρασης). Ορίζεται η μεταβολή της ενέργειας του συστήματος υπό σταθερή πίεση και θερμοκρασία].
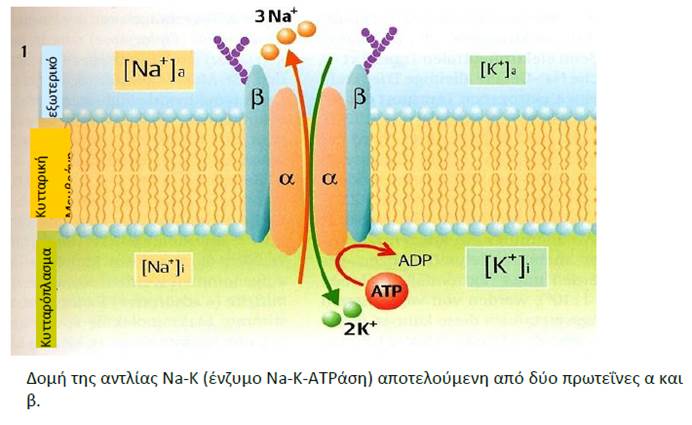
Για κάθε μόριο ATP που υδρολύεται, αντλούνται 3 Na+ προς τα έξω και 2 Κ+ προς τα μέσα. Το ένζυμο που συγκροτεί την αντλία ονομάζεται Κ-Νa-ΑΤΡάση. Ουσιαστικά η Κ-Νa-ΑΤΡάση και η αντλία ταυτίζονται.]
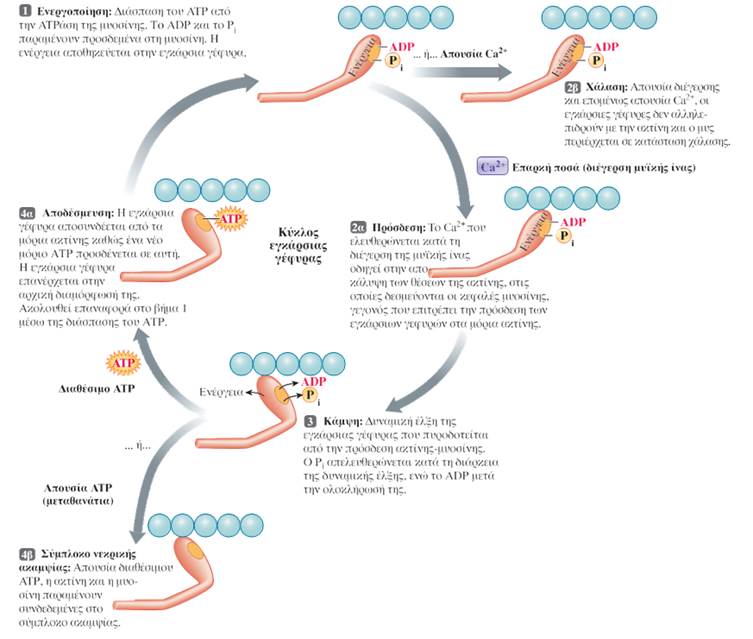
Η δεύτερη προσδένει τον φορέα ενέργειας τριφωσφορική αδενοσίνη (ATP) και τον υδρολύει σε διφωσφορική αδενοσίνη (ADP) και μία φωσφορική ομάδα (Pi), γεγονός που οδηγεί στην απελευθέρωση ενέργειας. Η διάσπαση της ATP λαμβάνει χώρα στην εγκάρσια γέφυρα της μυοσίνης πριν αυτή συνδεθεί με ένα μόριο ακτίνης. Η ADP και η φωσφορική ομάδα παραμένουν στενά προσδεμένες στο μόριο της μυοσίνης, ενώ η ενέργεια που απελευθερώνεται αποθηκεύεται στην εγκάρσια γέφυρα. Έτσι η μυοσίνη περιέρχεται σε μια κατάσταση «υψηλής ενέργειας».
Η δομή ενός λεπτού νηματίου:
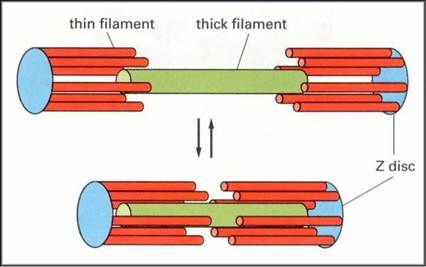
Οι δύο κεφαλές σε κάθε μόριο μυοσίνης λειτουργούν ανεξάρτητα η μία από την άλλη και ποτέ δε συνδέονται και οι δύο ταυτόχρονα στην ακτίνη. Κατά τη συστολή, τα λεπτά νημάτια σε κάθε πλευρά του σαρκομεριδίου ολισθαίνουν πάνω στα σταθερά παχέα νημάτια προς το κέντρο του, δηλαδή προς το κέντρο των ζωνών A. Καθώς τα λεπτά νημάτια ολισθαίνουν, έλκουν τις γραμμές Z στις οποίες είναι προσκολλημένα και έτσι τις φέρνουν πλησιέστερα τη μία στην άλλη, με συνέπεια το μήκος του σαρκομεριδίου να μειώνεται. Όλα τα σαρκομερίδια κατά μήκος της μυϊκής ίνας βραχύνονται ταυτόχρονα, με αποτέλεσμα να μειώνεται συνολικά το μήκος της. Ο παραπάνω τρόπος κίνησης των νηματίων αναφέρεται ως μηχανισμός διολίσθησης των νηματίων (sliding filament mechanism) και μέσω αυτού επιτυγχάνεται η μυϊκή συστολή.
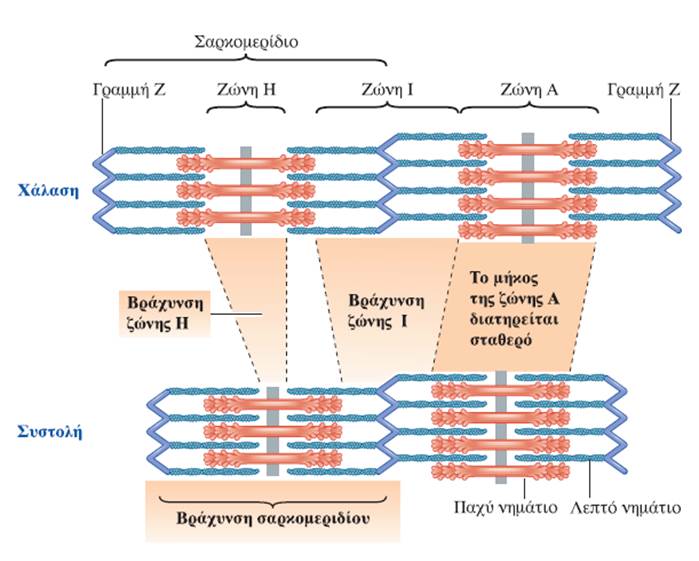
Μυϊκή Χάλαση και Συστολή:
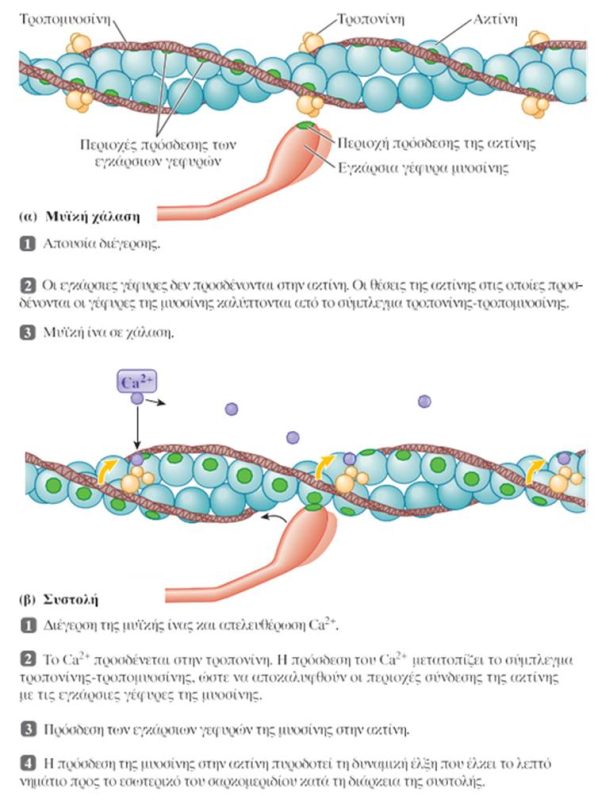
Η λειτουργία των εγκάρσιων γεφυρών:
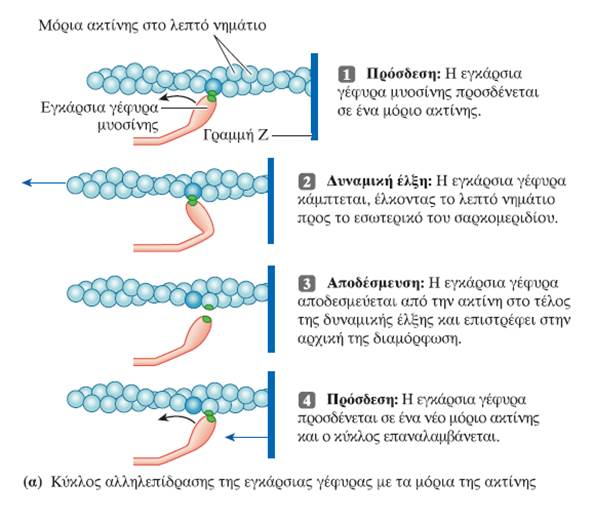
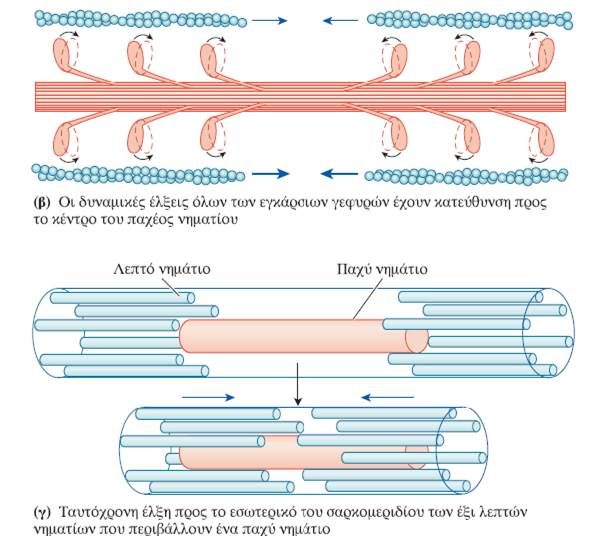
Ο μηχανισμός διολίσθησης των νηματίων κατά τη μυϊκή συστολή ενεργοποιείται από το Ca2+ που απελευθερώνεται από τους πλευρικούς σάκους του σαρκοπλασματικού δικτύου ως απόκριση στο δυναμικό ενέργειας που διαδίδεται μέσω των σωληναρίων Τ στο εσωτερικό της μυϊκής ίνας (σε κάθε περιοχή μετάβασης από τη ζώνη Α στη ζώνη Ι, η κυτταρική μεμβράνη της μυϊκής ίνας εγκολπώνεται και σχηματίζει ένα εγκάρσιο σωληνάριο ή σωληνάριο Τ (T tubule, Transverse tubule), το οποίο εκτείνεται κάθετα στην επιφάνεια της μυϊκής ίνας προς το κέντρο της. Επειδή η μεμβράνη του σωληναρίου Τ αποτελεί συνέχεια της επιφανειακής μεμβράνης της μυϊκής ίνας, ένα δυναμικό ενέργειας στην κυτταρική μεμβράνη διαδίδεται προς τα σωληνάρια Τ μεταδίδοντας ταχύτατα την επιφανειακή ηλεκτρική δραστηριότητα στο εσωτερικό της ίνας.
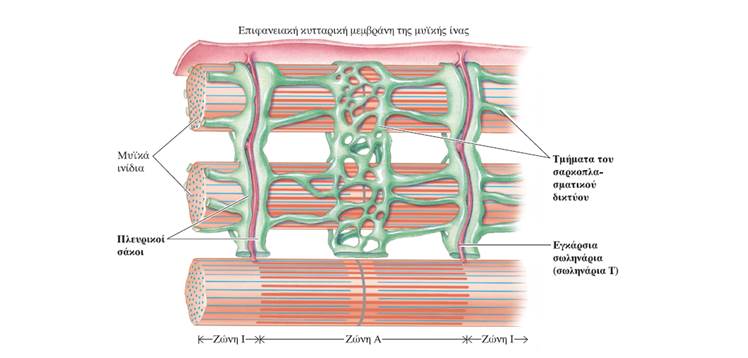
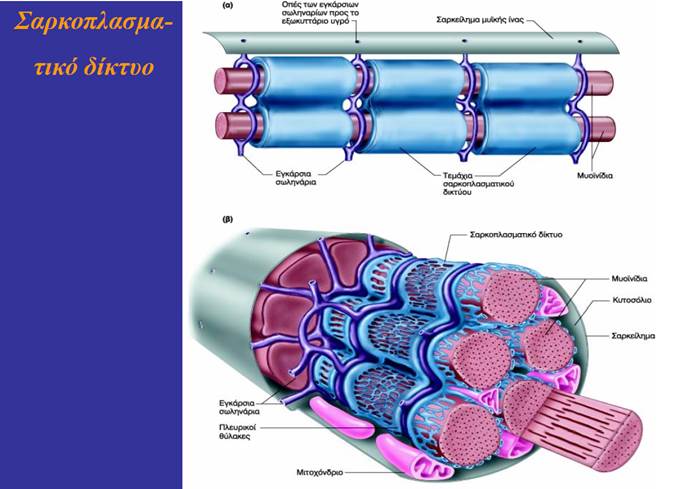
Το σαρκοπλασματικό δίκτυο (sarcoplasmic reticulum) είναι ένα τροποποιημένο ενδοπλασματικό δίκτυο. Πρόκειται για ένα μεμβρανικό δίκτυο δεξαμενών οι οποίες επικοινωνούν μεταξύ τους και περιβάλλουν κάθε μυϊκό ινίδιο. Τα μέρη που περιβάλλουν τις ζώνες Ι σχηματίζουν μια κάθετη αύλακα, που διαχωρίζει το καθένα από αυτά σε δύο πλευρικούς σάκους (lateral sacs), οι οποίοι είναι επίσης γνωστοί ως τελικές δεξαμενές (terminal cisternae). Την αύλακα αυτή διατρέχουν τα σωληνάρια Τ, διατηρώντας μια μικρή απόσταση από τους πλευρικούς σάκους. Οι πλευρικοί σάκοι αποθηκεύουν Ca2+. Η διάδοση του δυναμικού ενέργειας στα σωληνάρια Τ πυροδοτεί την απελευθέρωση Ca2+ από το σαρκοπλασματικό δίκτυο στο κυτταρόπλασμα της μυϊκής ίνας.).
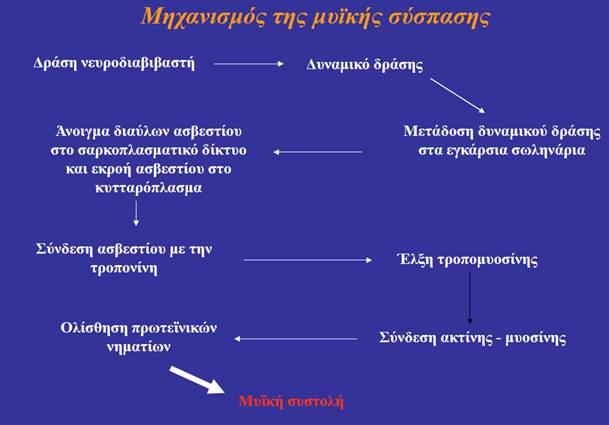
Ο όρος σύζευξη διέγερσης-συστολής (excitation-contraction coupling) αναφέρεται στα γεγονότα που συνδέουν τη διέγερση των μυϊκών ινιδίων (την παρουσία δηλαδή ενός δυναμικού ενέργειας στη μυϊκή ίνα) με τη μυϊκή συστολή (δηλαδή την αλληλουχία γεγονότων σύνδεσης-αποσύνδεσης των εγκάρσιων γεφυρών, που προκαλεί τη διολίσθηση των λεπτών νηματίων μέσω της οποίας επέρχεται η βράχυνση των σαρκομεριδίων). Το ασβέστιο είναι ο συνδετικός κρίκος μεταξύ της διέγερσης και της συστολής. Το Ca2+ που απελευθερώνεται δεσμεύεται στην τροπονίνη. Η πρόσδεση αυτή προκαλεί τη μετατόπιση της τροπομυοσίνης, ώστε να αποκαλυφθούν οι θέσεις των μορίων της ακτίνης, στις οποίες δεσμεύονται οι εγκάρσιες γέφυρες της μυοσίνης. Κατά την δυναμική έλξη, οι εγκάρσιες γέφυρες προσδένονται στην ακτίνη και ακολουθεί η έλξη των λεπτών νηματίων από τις δύο πλευρές του σαρκομεριδίου προς το μέσο του, που επιτυγχάνεται με την κάμψη των κεφαλών της μυοσίνης -όχι όλες ταυτόχρονα-. Η απαιτούμενη ενέργεια βρίσκεται αποθηκευμένη στην κεφαλή της μυοσίνης και προέρχεται από τη διάσπαση ATP που πραγματοποιεί η ATPάση της. Όταν ένα νέο μόριο ATP προσδεθεί στην εγκάρσια γέφυρα της μυοσίνης, τότε αυτή αποδεσμεύεται από την ακτίνη και επιστρέφει στην αρχική της θέση. Διαδοχικοί κύκλοι πρόσδεσης στην ακτίνη και κάμψης των εγκάρσιων γεφυρών έλκουν σταδιακά τα λεπτά νημάτια προς το εσωτερικό του σαρκομεριδίου.
Σύζευξη διέγερσης-συστολής και χάλαση μυών:
![]()
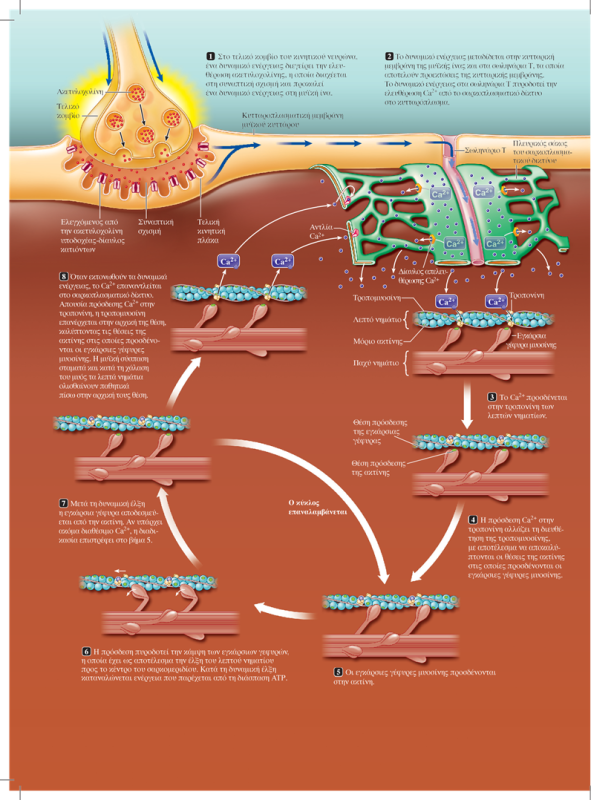
Όταν το δυναμικό ενέργειας εκτονωθεί, οι πλευρικοί σάκοι προσλαμβάνουν ενεργητικά το Ca2+, η τροπονίνη και η τροπομυοσίνη επανέρχονται στις θέσεις που αποκλείουν την πρόσδεση στα μόρια της ακτίνης των εγκάρσιων γεφυρών και επέρχεται η χάλαση του μυός.
Η κυκλική αλληλεπίδραση των εγκάρσιων γεφυρών με την ακτίνη:
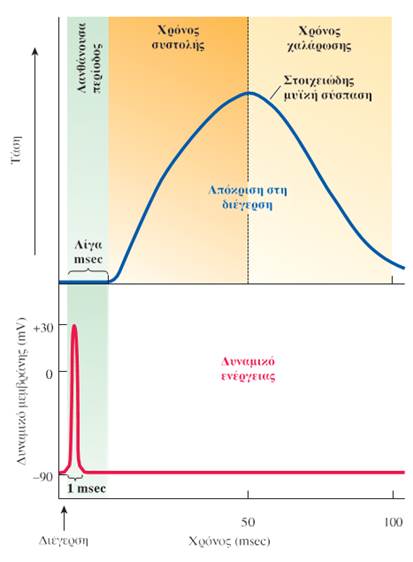
Η όλη διαδικασία της μυϊκής συστολής διαρκεί έως και 100 φορές περισσότερο από το δυναμικό ενέργειας. Στην πραγματικότητα, το δυναμικό ενέργειας έχει ήδη εκτονωθεί πριν ακόμα το συσταλτό σύστημα των μυών καταστεί λειτουργικό. Αυτή η χρονική καθυστέρηση λίγων msec ανάμεσα στη διέγερση και στην έναρξη της συστολής είναι γνωστή ως λανθάνουσα περίοδος (latent period). Χρόνος απαιτείται επίσης προκειμένου να αναπτυχθεί τάση (δύναμη συστολής) μέσα στη μυϊκή ίνα λόγω της ολίσθησης των λεπτών νηματίων πάνω στα παχέα νημάτια μέσω της δραστηριότητας των εγκάρσιων γεφυρών. Ο χρόνος από την έναρξη της συστολής έως την ανάπτυξη της μέγιστης τάσης ονομάζεται χρόνος συστολής (contraction time) και ποικίλλει από 15 έως 50 msec, ανάλογα με τον τύπο της μυϊκής ίνας. Η συστολή δε σταματά έως ότου οι πλευρικοί σάκοι επαναπροσλάβουν όλο το Ca2+ που απελευθερώθηκε ως απόκριση στο δυναμικό ενέργειας. Η επαναπρόσληψη του Ca2+ απαιτεί επίσης χρόνο. Καθώς το Ca2+ αντλείται στο εσωτερικό των πλευρικών σάκων, τα επίπεδά του στο κυτταρόπλασμα μειώνονται, με αποτέλεσμα οι προσδεμένες στην ακτίνη εγκάρσιες γέφυρες μυοσίνης και η συνολική δύναμη συστολής να μειώνονται. Ο χρόνος από το σημείο ανάπτυξης μέγιστης τάσης έως την ολοκλήρωση της χάλασης είναι γνωστός ως χρόνος χάλασης (relaxation time) και ποικίλλει από 15 έως 50 msec, ανάλογα πάλι με τον τύπο των μυϊκών ινών.
Μηχανική των σκελετικών μυών
Τα σαρκομερίδια αποτελούν το συσταλτό τμήμα του μυός και η μείωση του μήκους τους (η οποία προκαλείται από τον κύκλο των δυναμικών έλξεων των εγκάρσιων γεφυρών) έχει ως αποτέλεσμα την ανάπτυξη τάσης στον μυ. Η τάση (tension) αναπτύσσεται στο εσωτερικό των σαρκομεριδίων, τα οποία αποτελούν το συσταλτό τμήμα (contractile component) των μυών, ως αποτέλεσμα της ενεργοποίησης των εγκάρσιων γεφυρών μυοσίνης και της επακόλουθης διολίσθησης των λεπτών νηματίων.
Το άκρο του μυός που προσφύεται στο πιο ακίνητο (στατικό) τμήμα του σκελετού ονομάζεται έκφυση (origin), ενώ το άκρο που προσφύεται στο πιο κινητό τμήμα του σκελετού ονομάζεται κατάφυση (insertion). Για να μετακινηθεί το οστό στο οποίο ο μυς καταφύεται, αυτή η εσωτερική τάση που δημιουργεί η βράχυνση του συσταλτού τμήματος (των σαρκομεριδίων) μεταδίδεται στο οστό μέσω της διάτασης του εν σειρά ελαστικού στοιχείου του μυός, του τένοντα [οι τένοντες είναι αραιοκυτταρικές επιμήκεις δομές από καλά οργανωμένες ίνες κολλαγόνου και δεν περιέχουν μεγάλα αγγεία παρά μόνο λίγα τριχοειδή. Είναι ο μη συσταλτός συνδετικός ιστός που εκτείνεται πέρα από τα άκρα του μυός, σχηματίζοντας τους σκληρούς, κολλαγονικούς τένοντες (tendons) που συνδέουν τον μυ στα οστά.]. Το φορτίο είναι η αντιτιθέμενη δύναμη που ασκείται στον μυ από το βάρος ενός αντικειμένου.
Οι τρεις κύριοι τύποι συστολής είναι: (1) η ισοτονική (συστολή σταθερής τάσης), στην οποία το φορτίο παραμένει σταθερό ενώ ο μυς αλλάζει μήκος (βραχύνεται σε μια ομόκεντρη συστολή και επιμηκύνεται σε μια έκκεντρη συστολή), (2) ισοκινητική (συστολή σταθερής ταχύτητας), στην οποία η ταχύτητα της βράχυνσης παραμένει σταθερή καθώς ο μυς αλλάζει μήκος, και (3) ισομετρική (συστολή σταθερού μήκους), στην οποία το μήκος του μυός δεν αλλάζει καθώς αναπτύσσεται τάση.
[Είδη συστολής αναλυτικά:
Επειδή η λέξη συστολή εμπεριέχει την έννοια της βράχυνσης, μπορεί να προκαλέσει η φύση της μυϊκής συστολής αρχικά κάποια σύγχυση. Η μυϊκή συστολή συμβαίνει κάθε φορά που οι μυϊκές ίνες παράγουν τάση στα άκρα τους, κάτι που μπορεί να συμβεί όταν ένας μυς βραχύνεται, παραμένει στο ίδιο μήκος ή επιμηκύνεται.
Μειομετρική συστολή
Η μειομετρική συστολή συμβαίνει όταν η αναπτυσσόμενη τάση από τον μυ επαρκεί για να υπερνικήσει την αντίσταση και να μετακινήσει το τμήμα του σώματος της μιας πρόσφυσης προς το τμήμα της άλλης πρόσφυσης. Όταν ανυψώνεται το άνω άκρο προς τα πλάγια, βραχύνονται οι απαγωγοί μύες για να υπερνικήσουν την αντίσταση του άκρου. Ο μυς βραχύνεται και, όταν σταθεροποιηθεί η μια άκρη του, η άλλη έλκει το οστό προς εκεί και το στρέφει γύρω από τον άξονα της άρθρωσης.
Πλειομετρική συστολή
Όταν ένας μυς επιμηκύνεται αργά, καθώς ενδίδει σε μια εξωτερική δύναμη (όπως είναι η βαρύτητα), που είναι μεγαλύτερη από την συσταλτική δύναμη που μπορεί να ασκήσει, πρόκειται για μια πλειομετρική συστολή. Η χρήση του όρου επιμήκυνση μπορεί να θεωρηθεί παραπλανητική, γιατί στις περισσότερες περιπτώσεις δεν επιμηκύνεται πραγματικά ο μυς. Επανέρχεται απλώς από τη βραχυσμένη κατάστασή του στο φυσιολογικό μήκος ηρεμίας του. Οι απαγωγοί μύες του ώμου εκτελούν πλειομετρική συστολή, όταν κινείται το άνω άκρο από μια θέση απαγωγής προς τα κάτω. Στις περιπτώσεις όπου οι μύες συστέλλονται πλειομετρικά λειτουργούν ως «φρένο» ή δύναμη αντίστασης ενάντια στην κινητήρια δύναμη της βαρύτητας ή άλλη εξωτερική δύναμη. Όταν συστέλλονται οι μύες με αυτόν τον τρόπο, θεωρείται ότι παράγουν αρνητικό έργο.
Ισομετρική ή στατική συστολή
Η λέξη ισομετρική σημαίνει «ίσου μήκους». Η τάση του μυός σε μερική ή πλήρη συστολή, χωρίς κάποια ουσιαστική αλλαγή στο μήκος του, αποτελεί την ισομετρική συστολή. Η ισομετρική συστολή συμβαίνει συνήθως σε δύο διαφορετικές περιπτώσεις:
- Οι μύες που δρουν ανταγωνιστικά μεταξύ τους συστέλλονται με την ίδια δύναμη,
εξισορροπώνται και αλληλοεξουδετερώνονται. Το τμήμα, πάνω στο οποίο δρουν, συγκρατείται σταθερά στη θέση του, χωρίς να μετακινείται. Η σύσπαση του δικέφαλου για επίδειξη της γαστέρας του αποτελεί ένα παράδειγμα. Η σύσπαση του τρικέφαλου αποτρέπει τον αγκώνα από περαιτέρω κάμψη.
- Ένας μυς εκτελεί μερική ή μέγιστη συστολή ενάντια σε μια άλλη δύναμη, όπως είναι η έλξη της βαρύτητας, μια εξωτερική μηχανική ή μια άλλη μυϊκή δύναμη. Τέτοια παραδείγματα είναι όταν κρατάμε ένα βιβλίο με τεντωμένο χέρι, η διελκυστίνδα ανάμεσα σε ισάξιους αντιπάλους και η προσπάθεια να μετακινήσουμε ένα αντικείμενο που είναι πάρα πολύ βαρύ για τον σκοπό αυτό.
Ισοτονική συστολή
Η λέξη ισοτονικός σημαίνει «ίσης τάσης, έντασης». Η ισοτονική συστολή είναι μια συστολή, κατά την οποία η τάση παραμένει συνεχής, καθώς ο μυς επιμηκύνεται ή βραχύνεται. Χρησιμοποιείται συνήθως, αν και λανθασμένα, ως συνώνυμο για την μειομετρική ή την πλειομετρική συστολή. Οι όροι αυτοί δεν αναφέρονται στον βαθμό της τάσης αλλά στην αύξηση ή στην ελάττωση του μήκους.
Ισοκινητική συστολή
Η λέξη ισοκινητικός σημαίνει «ίσης ή ίδιας κίνησης». Με την χρήση ειδικού εξοπλισμού, είναι δυνατό να έχουμε μέγιστη μυϊκή προσπάθεια στην ίδια ταχύτητα για όλο το εύρος τροχιάς κίνησης του σχετικού μοχλού. Η αντίδραση των μυών σε μέγιστη συστολή προς την «προσαρμοζόμενη αντίσταση» του μηχανήματος ονομάζεται ισοκινητική συστολή.
Υπάρχει επίσης ένας άλλος τρόπος περιγραφής των μυϊκών συστολών, σύμφωνα με τον οποίο κατατάσσονται σε ομόκεντρες ή έκκεντρες. Στις ομόκεντρες ή σύγκεντρες συστολές (concentric contractions) ο μυς βραχύνεται, ενώ στις έκκεντρες συστολές (eccentric contractions) ο μυς επιμηκύνεται.]
Η ταχύτητα της βράχυνσης ενός μυός είναι αντιστρόφως ανάλογη του φορτίου. Το έργο που παράγεται από έναν συσπώμενο μυ ισούται με το γινόμενο του φορτίου επί την απόσταση κατά την οποία το μετακινεί. Η δύναμη (force) είναι η τάση που αναπτύσσει ο μυς προκειμένου να υπερνικηθεί το φορτίο (το βάρος του αντικειμένου). Το παραγόμενο έργο από έναν μυ που συσπάται αντιστοιχεί στο 0% έως 25% της συνολικής ενέργειας που αυτός καταναλώνει, ενώ η υπόλοιπη ενέργεια μετατρέπεται σε θερμότητα.
Πολλοί μύες είναι τμήματα συστημάτων μοχλών τα οποία επιτρέπουν η απόσταση και η ταχύτητα μετακίνησης των αντικείμενων να είναι πολλαπλάσιες της απόστασης και της ταχύτητας βράχυνσης του μυός. Ωστόσο, αυτός ο «πολλαπλασιασμός» γίνεται σε βάρος της δύναμης που ο μυς θα πρέπει να ασκήσει προκειμένου να μετακινήσει ένα φορτίο.
H μεταβολή της δύναμης που ασκείται από ολόκληρο τον μυ μπορεί να επιτευχθεί (1) με τη μεταβολή του αριθμού των κινητικών μονάδων (και κατ’ επέκταση του αριθμού των μυϊκών ινών) που συσπώνται στον μυ και (2) με τη διαβάθμιση της τάσης που αναπτύσσεται σε κάθε συσπώμενη μυϊκή ίνα.
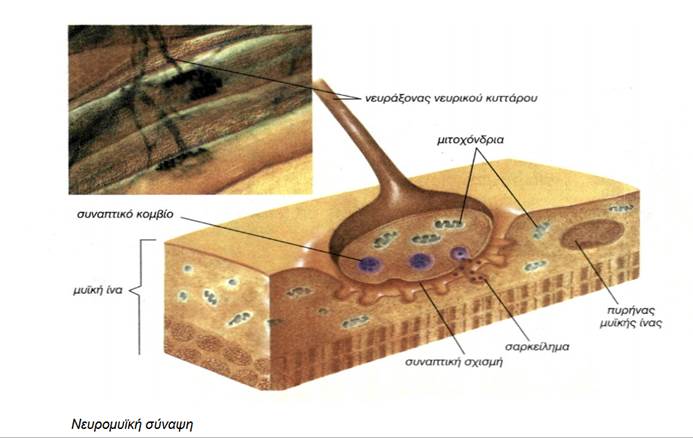
Κάθε συγκεκριμένος μυς νευρώνεται από διαφορετικό αριθμό κινητικών νευρώνων. Όταν ο κινητικός νευρώνας εισέρχεται στον μυ, διακλαδίζεται και κάθε τελική του απόληξη παρέχει νεύρωση σε μια μυϊκή ίνα. Κάθε κινητικός νευρώνας παρέχει νεύρωση σε έναν αριθμό μυϊκών ινών, ενώ κάθε μυϊκή ίνα νευρώνεται από έναν μόνο κινητικό νευρώνα. Όταν ένας κινητικός νευρώνας ενεργοποιείται, όλες οι μυϊκές ίνες που νευρώνει διεγείρονται, ώστε να συσπώνται ταυτόχρονα. Ένας νευρώνας δεν έχει ενδιάμεσο επίπεδο ενεργοποίησης. Είτε ενεργοποιείται, είτε όχι. Ο κινητικός νευρώνας και οι μυϊκές ίνες τις οποίες νευρώνει αποτελούν μια κινητική μονάδα (motor unit).
Οι μυϊκές ίνες που απαρτίζουν μια κινητική μονάδα είναι διεσπαρμένες σε ολόκληρο τον μυ, με αποτέλεσμα η ταυτόχρονη σύσπασή τους να οδηγεί σε μια ομαλά κατανεμημένη, αν και σχετικά αδύναμη, συστολή όλου του μυός. Κάθε μυς αποτελείται από ποικίλο αριθμό αναμειγμένων κινητικών μονάδων. Για μια αδύναμη σύσπαση του μυός ενεργοποιούνται μόνο μία ή λίγες από τις κινητικές του μονάδες.
Όσο περισσότερες κινητικές μονάδες στρατολογούνται και διεγείρονται ταυτόχρονα, τόσο αυξάνεται η συνολική δύναμη της συστολής του μυός. Το φαινόμενο αυτό είναι γνωστό ως στρατολόγηση κινητικών μονάδων (motor units recruitment).
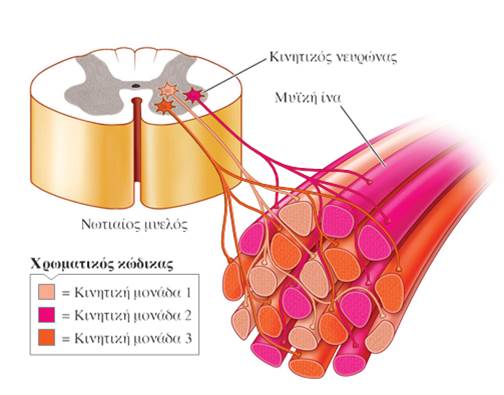
Ο αριθμός των μυϊκών ινών ενός μυός που συσπώνται εξαρτάται: (1) από τον αριθμό των κινητικών μονάδων που έχουν στρατολογηθεί (και κατ’ επέκταση τον αριθμό των κινητικών νευρώνων του μυός που είναι ενεργοί) και (2) από το μέγεθος των κινητικών αυτών μονάδων (δηλαδή τον αριθμό των μυϊκών ινών που ενεργοποιούνται ταυτόχρονα από έναν μεμονωμένο κινητικό νευρώνα).
Προκειμένου να καθυστερήσει ή να αποφευχθεί η κόπωση (fatigue – η αδυναμία διατήρησης της μυϊκής δύναμης σε ένα συγκεκριμένο επίπεδο) στην περίπτωση μυών που πρέπει να ασκούν για παρατεταμένα χρονικά διαστήματα δύναμη, λαμβάνει χώρα ασύγχρονη στρατολόγηση (asynchronous recruitment) των κινητικών μονάδων. Δηλαδή το σώμα εναλλάσσει τις κινητικές μονάδες που χρησιμοποιεί, ώστε να δίνει εκ περιτροπής την ευκαιρία στις κινητικές μονάδες να ξεκουράζονται.
Στις μέγιστες μυϊκές συστολές, καθώς όλες οι μυϊκές ίνες θα πρέπει να συσπαστούν, η εναλλαγή των κινητικών μονάδων ώστε να προληφθεί η κόπωση είναι εκ των πραγμάτων αδύνατη.
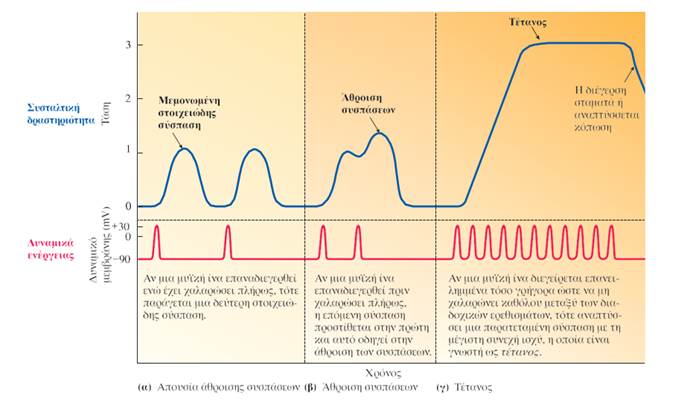
Ένα μεμονωμένο δυναμικό ενέργειας σε μια μυϊκή ίνα παράγει μια βραχεία, ασθενή συστολή που είναι γνωστή ως στοιχειώδης σύσπαση (twitch).
Δύο μεταβλητές που επηρεάζουν την τάση που αναπτύσσουν οι μυϊκές ίνες είναι: (1) η συχνότητα της διέγερσής τους, η οποία καθορίζει τον βαθμό της άθροισης των στοιχειωδών συσπάσεων, και (2) το μήκος τους πριν από την έναρξη της συστολής (σχέση μήκους-τάσης).
Ως άθροιση στοιχειωδών συσπάσεων αναφέρεται το φαινόμενο της αύξησης της τάσης που αναπτύσσει μια μυϊκή ίνα κατά την επαναλαμβανόμενη διέγερσή της, όταν το χρονικό διάστημα που μεσολαβεί ανάμεσα στα διαδοχικά δυναμικά ενέργειας δεν επαρκεί για την πλήρη χαλάρωσή της.
Η άθροιση των στοιχειωδών συσπάσεων οφείλεται στη διατήρηση των υψηλών επιπέδων Ca2+ στο κυτταρόπλασμα και στην παρατεταμένη διάταση του εν σειρά ελαστικού στοιχείου. Μετά την πάροδο της ανερέθιστης περιόδου ενός δυναμικού ενέργειας (κατά την οποία δεν είναι δυνατή η δημιουργία ενός νέου δυναμικού ενέργειας) και ενώ ακόμα διατηρείται ως έναν βαθμό η στοιχειώδης σύσπαση που αυτό προκάλεσε, η κυτταρική μεμβράνη του μυϊκού κυττάρου μπορεί να διεγερθεί από ένα νέο δυναμικό ενέργειας. Το γεγονός αυτό οδηγεί στην άθροιση της εναπομείνασας σύσπασης του προηγούμενου δυναμικού με αυτήν του νέου δυναμικού. Αν η συχνότητα διέγερσης της μυϊκής ίνας είναι τόσο υψηλή ώστε να μην έχει τη δυνατότητα να χαλαρώσει καθόλου ανάμεσα στα διαδοχικά ερεθίσματα, προκύπτει μια παρατεταμένη σύσπαση μέγιστης ισχύος που είναι γνωστή ως τέτανος.
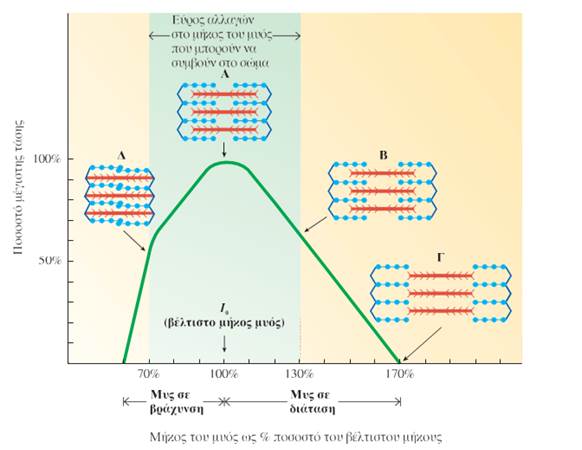
Η τάση που αναπτύσσει μια μυϊκή ίνα εξαρτάται επίσης από το μήκος της κατά την έναρξη της συστολής. Για κάθε μυ υπάρχει ένα βέλτιστο μήκος μυός στο οποίο μπορεί να αναπτυχθεί η μέγιστη τάση.
Στο βέλτιστο μήκος μυός αναπτύσσεται η μέγιστη τάση, επειδή ο βαθμός επικάλυψης των λεπτών νηματίων με τις περιοχές των παχέων νηματίων στις οποίες εντοπίζονται οι εγκάρσιες γέφυρες είναι ιδανικός και έτσι ο αριθμός των μορίων της ακτίνης και της μυοσίνης που μπορούν να αλληλεπιδράσουν μεταξύ τους είναι ο μέγιστος δυνατός. Σε μικρότερα ή μεγαλύτερα μήκη μυός αναπτύσσεται μικρότερη τάση.
Μεταβολισμός των σκελετικών μυών και τύποι μυϊκών ινών
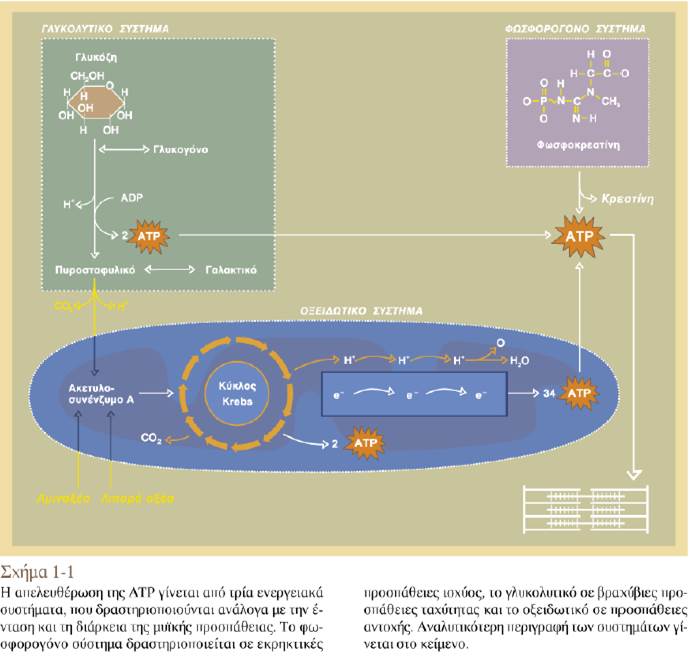
Τρεις μηχανισμοί εξασφαλίζουν το απαραίτητο ATP για τη συστολή και τη χάλαση των μυών: (1) η μεταφορά μιας υψηλής ενέργειας φωσφορικής ομάδας από την αποθηκευμένη φωσφορική κρεατίνη στην ADP, η οποία οδηγεί στον σχηματισμό των μορίων ATP που χρησιμοποιούνται πρώτα κατά την έναρξη της άσκησης, (2) η οξειδωτική φωσφορυλίωση, μια υψηλής απόδοσης διαδικασία παραγωγής ATP από τη διάσπαση των θρεπτικών συστατικών, η οποία χρησιμοποιείται όταν υπάρχει διαθέσιμο Ο2 και (3) η γλυκόλυση, κατά την οποία, όταν δεν υπάρχει Ο2, καταναλώνεται σε μεγάλες ποσότητες το αποθηκευμένο γλυκογόνο και συντίθεται ATP με παράλληλη παραγωγή γαλακτικού οξέος.
Οι τρεις κύριοι τύποι σκελετικών μυϊκών ινών με βάση τις διαφορές τους στο είδος των ενζυμικών αντιδράσεων που κυρίως βασίζονται για την παραγωγή ATP (οξειδωτική φωσφορυλίωση ή γλυκόλυση) και με βάση τις διαφορές τους στην ταχύτητα με την οποία διασπούν την ATP και ακολούθως συστέλλονται (βραδέως ή ταχέως) είναι οι ακόλουθες: (1) βραδείες οξειδωτικές ίνες, (2) ταχείες οξειδωτικές ίνες και (3) ταχείες γλυκολυτικές ίνες.
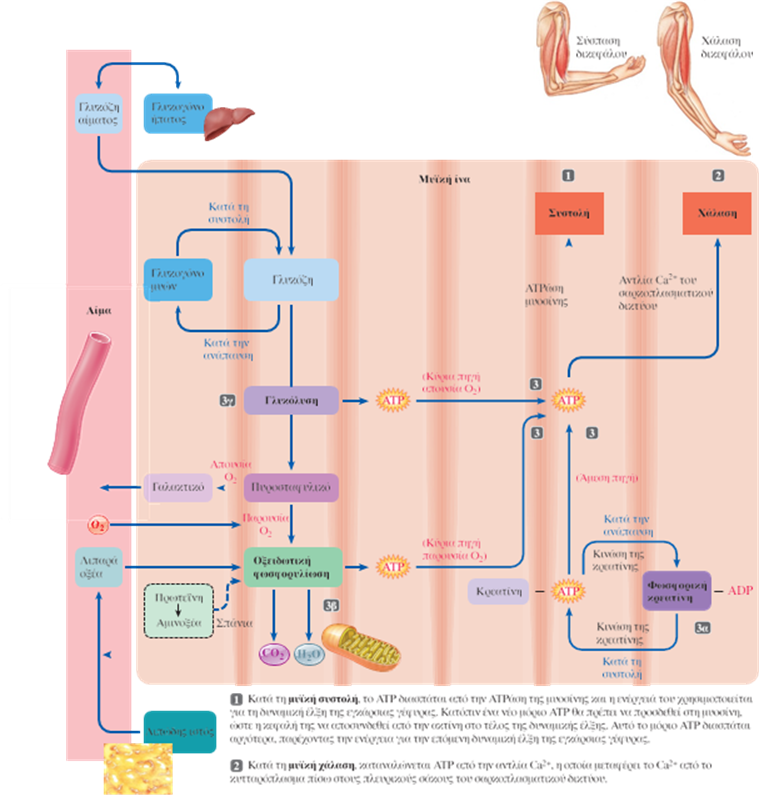
Στη διαδικασία διέγερσης, συστολής και χάλασης των μυών τέσσερα στάδια απαιτούν ATP:
- Η διάσπαση της ATP από την ATPάση της μυοσίνης παρέχει την ενέργεια για τη δυναμική έλξη της εγκάρσιας γέφυρας.
- Η πρόσδεση (όχι η διάσπαση) ενός νέου μορίου ATP στη μυοσίνη επιτρέπει την αποσύνδεση της εγκάρσιας γέφυρας από τα νημάτια της ακτίνης στο τέλος της δυναμικής έλξης, έτσι ώστε ο κύκλος να μπορεί να επαναληφθεί. Αυτή η ATP διασπάται αργότερα και παρέχει την ενέργεια για την επόμενη δυναμική έλξη της εγκάρσιας γέφυρας.
- Η ενεργητική μεταφορά του Ca2+ πίσω στους πλευρικούς σάκους του σαρκοπλασματικού δικτύου κατά τη διάρκεια της μυϊκής χάλασης καταναλώνει ενέργεια που προέρχεται από τη διάσπαση της ATP.
- Η ενεργητική μεταφορά του Na+ στο εξωκυτταρικό υγρό και του K+ στο ενδοκυτταρικό υγρό που ακολουθεί τη γένεση του δυναμικού ενέργειας στη μυϊκή ίνα διεκπεραιώνεται από την ATP-εξαρτώμενη αντλία K+-Na+.
Οι μυϊκές ίνες διαθέτουν εναλλακτικές οδούς παραγωγής ATP. Η ATP αποτελεί τη μοναδική πηγή ενέργειας που μπορεί να χρησιμοποιηθεί απευθείας για τις παραπάνω δραστηριότητες των μυϊκών κυττάρων. Επομένως η πραγματοποίηση της σύσπασης των σκελετικών μυών προϋποθέτει την παρουσία επαρκούς ποσότητας ATP. Ο μυϊκός ιστός αποθηκεύει μικρή μόνο ποσότητα ATP. Συνεπώς η διαθέσιμη ποσότητα προσχηματισμένου ATP δεν επαρκεί για την κάλυψη των αναγκών της μυϊκής συστολής. Ωστόσο, τα μυϊκά κύτταρα αντεπεξέρχονται στις ενεργειακές απαιτήσεις της μυϊκής συστολής χάρη σε τρεις μηχανισμούς παραγωγής περισσότερου ATP:
(1) τη μεταφορά μιας φωσφορικής ομάδας υψηλής ενέργειας από τη φωσφορική κρεατίνη στην ADP
(2) την οξειδωτική φωσφορυλίωση και
(3) τη γλυκόλυση
ΦΩΣΦΟΡΙΚΗ ΚΡΕΑΤΙΝΗ
Η φωσφορική κρεατίνη (creatine phosphate) είναι η πρώτη αποθήκη ενέργειας που κινητοποιείται κατά την έναρξη της συστολής. Η υψηλής ενέργειας φωσφορική ομάδα της φωσφορικής κρεατίνης μπορεί να μεταφερθεί απευθείας στην ADP και έτσι να σχηματίσει ATP. Όπως ακριβώς εκλύεται ενέργεια από τη διάσπαση της τελικής φωσφορικής ομάδας της ATP, έτσι ελευθερώνεται ενέργεια και από τη διάσπαση του δεσμού μεταξύ της φωσφορικής ομάδας και της κρεατίνης. Η ενέργεια που ελευθερώνεται από την υδρόλυση της φωσφορικής κρεατίνης, μαζί με τη φωσφορική ομάδα, μπορεί να μεταφερθεί κατευθείαν στην ADP, ώστε να σχηματιστεί ATP. Αυτή η αντίδραση καταλύεται από ένα μυϊκό ένζυμο, την κινάση της κρεατίνης (creatine kinase), και είναι αμφίδρομη. Δηλαδή η ενέργεια και η φωσφορική ομάδα μπορεί να μεταφερθούν και από την ATP στην κρεατίνη, ώστε να σχηματιστεί φωσφορική κρεατίνη.
Φωσφορική κρεατίνη + ADP ← κινάση της κρεατίνης → κρεατίνη + ATP
Όταν το μυϊκό κύτταρο βρίσκεται σε ηρεμία, η συγκέντρωση της ATP σε αυτό αυξάνεται, οπότε σύμφωνα με τον νόμο δράσης των μαζών ευνοείται η μεταφορά της φωσφορικής ομάδας υψηλής ενέργειας από την ATP στην κρεατίνη, ώστε να σχηματιστεί φωσφορική κρεατίνη.
Αντίθετα, κατά την έναρξη της μυϊκής σύσπασης, όταν η ATPάση της μυοσίνης καταναλώνει γρήγορα τα περιορισμένα αποθέματα ATP, η μείωση στη συγκέντρωση της ATP ευνοεί τη μεταφορά της φωσφορικής ομάδας υψηλής ενέργειας από την αποθηκευμένη φωσφορική κρεατίνη στην ADP, ώστε να σχηματιστεί νέο ATP.
Οι μύες σε ηρεμία περιέχουν περίπου 5 φορές περισσότερη φωσφορική κρεατίνη απ’ ό,τι ATP. Έτσι το μεγαλύτερο μέρος της ενέργειας που αποθηκεύουν βρίσκεται στη μορφή φωσφορικής κρεατίνης. Η μεταφορά της φωσφορικής ομάδας από τη φωσφορική κρεατίνη στο ATP πραγματοποιείται ταχύτατα (σε κλάσματα του δευτερολέπτου), καθώς απαιτείται μία μόνο ενζυμική αντίδραση.
Η φωσφορική κρεατίνη είναι η πρώτη πηγή ενέργειας που αξιοποιείται για την παραγωγή επιπλέον ATP κατά την έναρξη της σωματικής άσκησης.
Τα επίπεδα ATP στους μυς παραμένουν αρκετά σταθερά στην αρχή της μυϊκής δραστηριότητας, χάρη στην αξιοποίηση της αποθηκευμένης φωσφορικής κρεατίνης, η οποία όμως εξαντλείται γρήγορα. Η μικρής χρονικής διάρκειας έντονη σωματική άσκηση (για παράδειγμα, ψηλά άλματα, γρήγορο τρέξιμο ή άρση βαρών) καλύπτεται ενεργειακά κυρίως από την ATP που παράγεται από τη διάσπαση της φωσφορικής κρεατίνης. Τα αποθέματα φωσφορικής κρεατίνης τυπικά εξασφαλίζουν την ενέργεια που χρειάζεται για το πρώτο λεπτό (ή ακόμα λιγότερο) της άσκησης.
Φυσιολογικά προσλαμβάνουμε την κρεατίνη με την τροφή μας. Ωστόσο, ορισμένοι αθλητές, ελπίζοντας να αποκτήσουν αγωνιστικό πλεονέκτημα, λαμβάνουν διατροφικά συμπληρώματα κρεατίνης, με σκοπό να βελτιώσουν την επίδοσή τους σε δραστηριότητες που απαιτούν έντονη μυϊκή προσπάθεια διάρκειας μικρότερης του ενός λεπτού. Η τροφοδότηση των μυών τους με περισσότερη κρεατίνη οδηγεί σε μεγαλύτερα αποθέματα φωσφορικής κρεατίνης και αυτό με τη σειρά του μεταφράζεται σε μικρή βελτίωση της απόδοσης σε αθλήματα που απαιτούν κατανάλωση μεγάλων ποσοτήτων ενέργειας σε μικρό χρόνο. Τα συμπληρώματα κρεατίνης όμως θα πρέπει να λαμβάνονται με προσοχή λόγω του ότι οι μακροχρόνιες επιπτώσεις τους στην υγεία δεν είναι γνωστές. Επιπλέον, μεγαλύτερα αποθέματα φωσφορικής κρεατίνης δεν προσφέρουν κανένα όφελος σε σωματικές δραστηριότητες μεγαλύτερης διάρκειας, καθώς αυτές βασίζονται σε άλλους μηχανισμούς που εξασφαλίζουν την παροχή ενέργειας για μεγαλύτερα χρονικά διαστήματα.
ΟΞΕΙΔΩΤΙΚΗ ΦΩΣΦΟΡΥΛΙΩΣΗ
Η οξειδωτική φωσφορυλίωση είναι μια διαδικασία πολλών βημάτων από την οποία παράγεται ATP με χαμηλότερο ρυθμό σε σχέση με την παραγωγή ATP μέσω της φωσφορικής κρεατίνης ή της γλυκόλυσης.
Η οξειδωτική φωσφορυλίωση λαμβάνει χώρα στα μιτοχόνδρια, εφόσον υπάρχει επαρκής ποσότητα Ο2. Το Ο2 είναι απαραίτητο για τη λειτουργία της αλυσίδας μεταφοράς ηλεκτρονίων, μέσω της οποίας η χημική ενέργεια που απελευθερώνεται από τη διάσπαση των θρεπτικών συστατικών χρησιμοποιείται για την παραγωγή ATP.
Η σύνθεση της ATP πραγματοποιείται από την ATP συνθετάση κατά τη χημειώσμωση. Τα κύρια θρεπτικά συστατικά που χρησιμοποιούνται σε αυτή την οδό είναι η γλυκόζη και τα λιπαρά οξέα, ανάλογα με την ένταση και τη διάρκεια της σωματικής δραστηριότητας.
Αν και η οξειδωτική φωσφορυλίωση έχει υψηλή απόδοση σε ATP καθώς παράγονται 32 μόρια ATP για κάθε μόριο γλυκόζης που διασπάται, είναι σχετικά αργή λόγω του μεγάλου αριθμού των ενζυμικών βημάτων που περιλαμβάνει.
Τα μυϊκά κύτταρα μπορούν να παράγουν από την οξειδωτική φωσφορυλίωση επαρκείς ποσότητες ATP, ώστε να ανταποκρίνονται για παρατεταμένες περιόδους σε μέτριες ενεργειακές ανάγκες των μυών, όπως αυτές που δημιουργούνται κατά την ήπια σωματική άσκηση (για παράδειγμα, το περπάτημα) ή τη σωματική άσκηση μέσης έντασης (για παράδειγμα, το κολύμπι ή το τζόκινγκ).
Προκειμένου να συντηρήσουν την οξειδωτική φωσφορυλίωση, οι μύες χρειάζονται συνεχή τροφοδοσία με επαρκείς ποσότητες Ο2 και θρεπτικών συστατικών. Με την οξειδωτική φωσφορυλίωση υποστηρίζεται ενεργειακά η λεγόμενη αερόβια άσκηση (aerobic exercise) ή άσκηση αντοχής (endurance-type exercise). Το Ο2 που απαιτείται για την οξειδωτική φωσφορυλίωση μεταφέρεται στους μυς με την κυκλοφορία του αίματος. Οι μύες εξασφαλίζουν την αυξημένη ποσότητα Ο2 που χρειάζονται κατά τη διάρκεια της άσκησης με διάφορους μηχανισμούς (οι οποίοι συζητούνται αναλυτικότερα παρακάτω):
Η ταχύτερη και βαθύτερη αναπνοή αυξάνει τη συγκέντρωση του Ο2 στο αρτηριακό αίμα, η καρδιά συστέλλεται με αυξημένη συχνότητα και δύναμη ώστε να αντλήσει περισσότερο οξυγονωμένο αίμα στους ιστούς, περισσότερο αίμα τροφοδοτεί τους ασκούμενους μυς λόγω της διαστολής των αιμοφόρων αγγείων που τους αιματώνουν και η αιμοσφαιρίνη, ο κύριος μεταφορέας οξυγόνου στο αίμα, ελευθερώνει περισσότερο Ο2 στους ασκούμενους μυς.
Επιπλέον, ορισμένοι τύποι μυϊκών ινών διαθέτουν μεγάλες ποσότητες μυοσφαιρίνης (myoglobin), μιας πρωτεΐνης που μοιάζει με την αιμοσφαιρίνη. Η μυοσφαιρίνη μπορεί να αποθηκεύσει μικρή μόνο ποσότητα Ο2, αλλά το πιο σημαντικό είναι ότι αυξάνει τον ρυθμό μεταφοράς Ο2 από το αίμα στις μυϊκές ίνες.
Η γλυκόζη και τα λιπαρά οξέα, που προέρχονται από τη διατροφή, μεταφέρονται στα μυϊκά κύτταρα με την κυκλοφορία του αίματος. Επιπλέον, τα μυϊκά κύτταρα μπορούν να αποθηκεύσουν περιορισμένες ποσότητες γλυκόζης στη μορφή γλυκογόνου (αλυσίδες γλυκόζης). Το ήπαρ επίσης έχει την ικανότητα, έως έναν βαθμό, να αποθηκεύει την περίσσεια των προσλαμβανόμενων υδατανθράκων στη μορφή γλυκογόνου, το οποίο, μεταξύ των γευμάτων, μπορεί να διασπάται σε γλυκόζη και να αποδίδεται στην κυκλοφορία του αίματος. Η φόρτιση με υδατάνθρακες, η λήψη δηλαδή αυξημένων ποσοτήτων υδατανθράκων πριν από ένα αγώνισμα, είναι μια τακτική, η οποία χρησιμοποιείται από μερικούς αθλητές που ελπίζουν ότι θα βελτιώσουν έτσι την απόδοσή τους σε αθλήματα αντοχής όπως ο μαραθώνιος. Όμως, όταν οι αποθήκες γλυκογόνου των μυών και του ήπατος κορεστούν, οι επιπλέον ποσότητες υδατανθράκων (όπως και οποιοδήποτε άλλο θρεπτικό συστατικό πλούσιο σε ενέργεια) μετατρέπονται σε σωματικό λίπος.
ΓΛΥΚΟΛΥΣΗ
Υπάρχουν όρια στην ποσότητα του Ο2 που μπορεί να μεταφερθεί στους μυς, τα οποία τίθενται από το κυκλοφορικό και το αναπνευστικό σύστημα – οι πνεύμονες και η καρδιά μπορούν να προσλάβουν και να μεταφέρουν στους ασκούμενους μυς Ο2 μέχρι ενός ορίου. Επιπλέον, όταν ένας μυς συσπάται με σχεδόν μέγιστη δύναμη, λόγω της ισχυρής συστολής τα αιμοφόρα αγγεία που τον αιματώνουν πιέζονται, γεγονός που περιορίζει σημαντικά την τροφοδοσία του σε Ο2. Ακόμα όμως και αν το Ο2 δεν αποτελεί περιοριστικό παράγοντα, ο σχετικά αργός ρυθμός παραγωγής ATP μέσω της οξειδωτικής φωσφορυλίωσης πιθανόν να μην είναι ικανός να παρέχει αρκετή ενέργεια στους μυς κατά τη διάρκεια έντονης άσκησης.
Η κατανάλωση ενέργειας από έναν σκελετικό μυ μπορεί να αυξηθεί έως και 100 φορές όταν αυτός μεταβαίνει από την ηρεμία σε άσκηση υψηλής έντασης. Όταν η παροχή Ο2 στους μυς ή η οξειδωτική φωσφορυλίωση δεν επαρκεί για να καλύψει τις ανάγκες σε ATP, καθώς η ένταση της άσκησης αυξάνεται, οι μυϊκές ίνες εξαρτώνται σταδιακά ολοένα και περισσότερο από τη γλυκόλυση για την παραγωγή ATP.
Οι χημικές αντιδράσεις της γλυκόλυσης παράγουν προϊόντα τα οποία εισέρχονται τελικά στην οξειδωτική φωσφορυλίωση, αλλά η γλυκόλυση μπορεί επίσης να λαμβάνει χώρα ανεξάρτητα από την περαιτέρω επεξεργασία των προϊόντων της μέσω της οξειδωτικής φωσφορυλίωσης.
Κατά τη διάρκεια της γλυκόλυσης, κάθε μόριο γλυκόζης διασπάται σε δύο μόρια πυροσταφυλικού οξέος, παράγοντας δύο μόρια ATP. Το πυροσταφυλικό οξύ μπορεί στη συνέχεια να εισέλθει στην οξειδωτική φωσφορυλίωση και να διασπαστεί περαιτέρω εξασφαλίζοντας περισσότερη ενέργεια.
Η γλυκόλυση έχει δύο πλεονεκτήματα σε σχέση με την οξειδωτική φωσφορυλίωση: (1) παράγει ATP απουσία Ο2 (λαμβάνει χώρα αναερόβια ) και (2) είναι πολύ ταχύτερη από την οξειδωτική φωσφορυλίωση.
Αν και η γλυκόλυση από μόνη της παράγει σημαντικά χαμηλότερη ποσότητα ATP από κάθε μόριο γλυκόζης που διασπάται, λόγω της μεγαλύτερης ταχύτητάς της, ο ρυθμός με τον οποίο παράγει ATP μπορεί να ξεπερνά αυτόν της οξειδωτικής φωσφορυλίωσης.
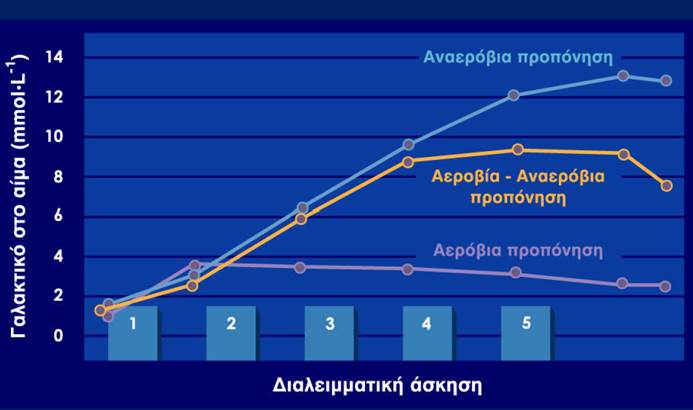
Με τη γλυκόλυση υποστηρίζεται ενεργειακά η λεγόμενη αναερόβια άσκηση (anaerobic exercise) ή άσκηση υψηλής έντασης (high-intensity exercise).
ΠΑΡΑΓΩΓΗ ΓΑΛΑΚΤΙΚΟΥ ΟΞΕΟΣ
Η γλυκόλυση παρέχει έναν αναερόβιο βιοχημικό μηχανισμό για την ενεργειακή υποστήριξη της έντονης σωματικής άσκησης όταν η παροχή Ο2 ή η παραγωγικότητα σε ATP της οξειδωτικής φωσφορυλίωσης δεν μπορεί να καλύψει τις ενεργειακές ανάγκες των μυών. Ωστόσο, η χρησιμοποίησή της έχει δύο συνέπειες.
Πρώτον, μεγάλα ποσά γλυκόζης θα πρέπει να διασπαστούν, λόγω του ότι η γλυκόλυση είναι λιγότερο αποτελεσματική σε σχέση με την οξειδωτική φωσφορυλίωση στη μετατροπή της χημικής ενέργειας των θρεπτικών συστατικών σε ενέργεια της ATP (η γλυκόλυση αποδίδει καθαρά δύο μόρια ATP για κάθε μόριο γλυκόζης που διασπάται, ενώ η οξειδωτική φωσφορυλίωση αποδίδει 32 μόρια ATP για κάθε μόριο γλυκόζης). Τα μυϊκά κύτταρα μπορούν να αποθηκεύσουν περιορισμένες ποσότητες γλυκόζης στη μορφή γλυκογόνου και η γλυκόλυση εξαντλεί τα αποθέματα του γλυκογόνου ταχύτατα.
Δεύτερον, όταν το τελικό προϊόν της γλυκόλυσης, το πυροσταφυλικό οξύ, δεν μπορεί να διασπαστεί περαιτέρω με την οξειδωτική φωσφορυλίωση, μετατρέπεται σε γαλακτικό οξύ (lactic acid), η συσσώρευση του οποίου συμβάλλει στη δημιουργία του μυϊκού πόνου που προκαλείται κατά τη διάρκεια της έντονης σωματικής άσκησης.
Ωστόσο, ο πόνος και η δυσκαμψία των μυών που εμφανίζονται την επόμενη μέρα της έντονης σωματικής άσκησης είναι πιθανό να οφείλονται σε αντιστρεπτές δομικές βλάβες των μυών. Επιπλέον, το γαλακτικό οξύ που παράγεται από τους μυς μεταφέρεται στο αίμα και είναι υπεύθυνο για τη μεταβολική οξέωση που παρατηρείται κατά την έντονη σωματική άσκηση. Συνεπώς οι αναερόβιες ασκήσεις υψηλής έντασης μπορεί να διατηρηθούν για μικρό μόνο χρονικό διάστημα, σε αντίθεση με τις αερόβιες ασκήσεις αντοχής, στις οποίες ένας αθλούμενος μπορεί να ασκείται συνεχόμενα για μεγάλο χρονικό διάστημα.
Η κόπωση μπορεί να είναι μυϊκής ή κεντρικής αιτιολογίας. Η συσταλτική δραστηριότητα σε έναν σκελετικό μυ δεν μπορεί να διατηρείται στο διηνεκές σε υψηλά επίπεδα. Αντιθέτως, η τάση που αναπτύσσεται από τον μυ μειώνεται καθώς αυτός εμφανίζει κόπωση. Υπάρχουν δύο τύποι κόπωσης: η μυϊκή και η κεντρική κόπωση.
Η μυϊκή κόπωση (muscle fatigue) συμβαίνει όταν ένας ασκούμενος μυς δεν μπορεί πλέον να ανταποκριθεί στη διέγερση με την ίδιας ισχύος συστολή. Η μυϊκή κόπωση αποτελεί έναν αμυντικό μηχανισμό που προστατεύει τους μυς, ώστε να μην υπερβούν το σημείο μετά το οποίο δε θα είναι πλέον ικανοί να παράγουν ATP. Η αδυναμία παραγωγής ATP θα κατέληγε σε νεκρική ακαμψία (που προφανώς δεν είναι μια επιθυμητή κατάληξη της σωματικής άσκησης). Οι αιτίες της μυϊκής κόπωσης δεν είναι σαφείς, ωστόσο κύριοι παράγοντες που εμπλέκονται σε αυτή θεωρούνται οι ακόλουθοι:
-Η τοπική αύξηση των επιπέδων του ανόργανου φωσφόρου από τη διάσπαση της ATP θεωρείται η βασική αιτία της μυϊκής κόπωσης. Τα αυξημένα επίπεδα Pi μειώνουν τη δύναμη συστολής επιδρώντας στη δυναμική έλξη των κεφαλών της μυοσίνης. Επιπλέον, τα αυξημένα επίπεδα Pi φαίνεται ότι ελαττώνουν την ανταπόκριση της τροπονίνης στο Ca2+ και μειώνουν την ποσότητα του Ca2+ που ελευθερώνεται από τους πλευρικούς σάκους.
-Η εξάντληση των ενεργειακών αποθεμάτων γλυκογόνου μπορεί επίσης να οδηγήσει σε μυϊκή κόπωση μετά από εξαντλητική άσκηση. Ο χρόνος εμφάνισης της κόπωσης ποικίλλει ανάλογα με τον τύπο της μυϊκής ίνας (κάποιες μυϊκές ίνες είναι περισσότερο ανθεκτικές στην κόπωση σε σχέση με άλλες) και την ένταση της άσκησης (δραστηριότητες υψηλής έντασης επιφέρουν ταχύτερα κόπωση).
Η κεντρική κόπωση (central fatigue) συμβαίνει όταν το κεντρικό νευρικό σύστημα (ΚΝΣ) δεν ενεργοποιεί πλέον επαρκώς τους κινητικούς νευρώνες που νευρώνουν τους συσπώμενους μυς. Στην περίπτωση αυτή, το άτομο επιβραδύνει ή σταματά τη σωματική άσκηση παρόλο που οι μύες του είναι ακόμα σε θέση να παράγουν έργο. Τα αίτια της κεντρικής κόπωσης είναι συχνά ψυχολογικά. Η κεντρική κόπωση μπορεί να πηγάζει από τη δυσφορία που προκαλεί η έντονη σωματική άσκηση. Προκειμένου να υπερνικήσει κάποιος τη δυσφορία αυτή και να συνεχίσει την άσκηση, χρειάζεται ισχυρή θέληση, η οποία συνήθως συνδέεται με κάποιο κίνητρο, για παράδειγμα την επιθυμία να διακριθούμε σε κάποιο αγώνισμα. Σε λιγότερο έντονες σωματικές δραστηριότητες, η κεντρική κόπωση μπορεί να οφείλεται στην ανία που προκαλεί μια μονότονη ενασχόληση (για παράδειγμα, η εργασία σε μια γραμμή παραγωγής) ή στη γενικότερη κούραση (για παράδειγμα, λόγω έλλειψης ύπνου) και έτσι να προκαλείται μείωση της απόδοσης. Οι μηχανισμοί που εμπλέκονται στην κεντρική κόπωση είναι σε μικρό μόνο βαθμό κατανοητοί. Σε κάποιες περιπτώσεις, η κεντρική κόπωση μπορεί να οφείλεται σε αυξημένα επίπεδα του νευροδιαβιβαστή σεροτονίνη και του αμινοξέος τρυπτοφάνη, που είναι η πρόδρομη ουσία της σεροτονίνης στον εγκέφαλο.
Η ανάκαμψη από την άσκηση απαιτεί αυξημένη κατανάλωση Ο2. Ένα άτομο συνεχίζει να αναπνέει βαθιά και γρήγορα για κάποιο χρονικό διάστημα μετά την άσκηση. Η ανάγκη αυξημένης πρόσληψης Ο2 κατά τη διάρκεια της ανάκαμψης από την άσκηση ονομάζεται αυξημένη κατανάλωση οξυγόνου μετά την άσκηση (EPOC, Excess Postexercise Oxygen Consumption) και οφείλεται σε ποικίλους παράγοντες.
Ο πιο γνωστός από αυτούς είναι η αναπλήρωση του ελλείμματος οξυγόνου (oxygen deficit) που δημιουργείται στη διάρκεια της άσκησης και έχει ως συνέπεια η μυϊκή σύσπαση να υποστηρίζεται ενεργειακά από ATP που παράγεται με μη οξειδωτικούς μηχανισμούς, όπως μέσω της φωσφορικής κρεατίνης και της αναερόβιας γλυκόλυσης.
[Χρέος Οξυγόνου (oxygen deficit)
Το Χρέος Ο2 δημιουργείται όταν οι απαιτήσεις για κατανάλωση Ο2 υπερβαίνουν την παροχή σε Ο2. Η ενέργεια παράγεται μέσω αναερόβιων μηχανισμών. Το μέγεθος και η διάρκεια του χρέους Ο2 εξαρτάται από την διάρκεια, την ένταση και το είδος της άσκησης. Κατά την επιτέλεση βαριάς μυϊκής εργασίας, παρά τη μεγάλη αύξηση του ποσού του οξυγόνου που μεταφέρεται στους μυς, αυτό δεν επαρκεί για την απελευθέρωση όλης της ενέργειας που απαιτείται για την επιτέλεση αυτού του έργου. Στις περιπτώσεις αυτές, ένα μέρος από το γλυκογόνο των μυών διασπάται αναερόβια, δηλαδή χωρίς οξειδωτικές διεργασίες, αποδίδοντας επιπρόσθετη ενέργεια, με αποτέλεσμα την παραγωγή γαλακτικού οξέος. Δηλαδή στις περιπτώσεις αυτές δημιουργείται στον οργανισμό μια κατάσταση που μπορεί να χαρακτηρίζεται ως “χρέος οξυγόνου”. Το έργο που επιτελείται με ενέργεια που απελευθερώνεται με αυτό τον τρόπο χαρακτηρίζεται ως αναερόβιο έργο. Μετά το τέλος μιας τέτοιας βαριάς μυϊκής εργασίας, το άτομο εξακολουθεί να αναπνέει έντονα και να προσλαμβάνει και να καταναλίσκει οξυγόνο με ρυθμό μεγαλύτερο από τον συνήθη ρυθμό σε ηρεμία, για αρκετά λεπτά, μέχρις ότου οι μεταβολικές διεργασίες στο σώμα του να επανέλθουν στον πριν από την μυϊκή εργασία ρυθμό. Δηλαδή, κατά το χρονικό αυτό διάστημα πραγματοποιείται η “αποπληρωμή του χρέους οξυγόνου”.]
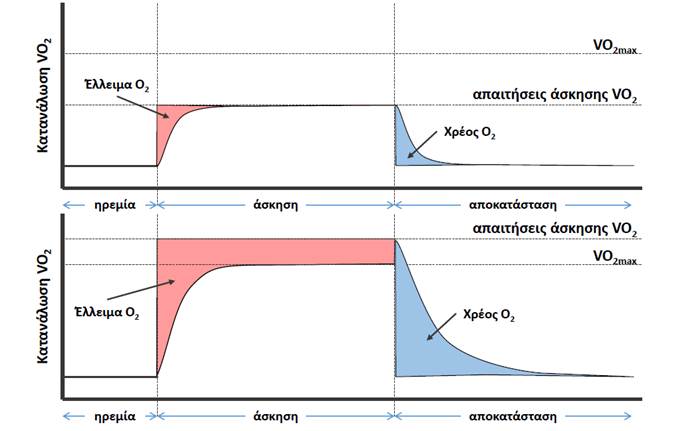
Κατά τη διάρκεια της άσκησης μειώνονται τα επίπεδα της φωσφορικής κρεατίνης των μυών, αθροίζεται γαλακτικό οξύ και μπορεί επίσης να μειώνεται η ποσότητα του αποθηκευμένου γλυκογόνου. Ο βαθμός στον οποίο παρατηρούνται τα παραπάνω εξαρτάται από την ένταση και τη διάρκεια της άσκησης.
Για την αποκατάσταση των ενεργειακών αποθεμάτων απαιτείται Ο2. Κατά τη διάρκεια της περιόδου ανάκαμψης παράγεται με οξειδωτική φωσφορυλίωση και αποθηκεύεται ATP, μέσω της κατανάλωσης των αυξημένων επιπέδων Ο2, με το οποίο τροφοδοτείται το σώμα μετά το τέλος της άσκησης χάρη στη βαθιά και γρήγορη αναπνοή.
Η μεγαλύτερη ποσότητα από την αναγεννώμενη ATP χρησιμοποιείται για την επανασύνθεση της φωσφορικής κρεατίνης ώστε να αποκατασταθούν τα αποθέματά της, γεγονός που μπορεί να επιτευχθεί σε χρονικό διάστημα λίγων λεπτών.
Το γαλακτικό οξύ που έχει συσσωρευτεί κατά την άσκηση μετατρέπεται σε πυροσταφυλικό οξύ, μέρος του οποίου χρησιμοποιείται για να παραχθούν νέα μόρια ATP μέσω οξειδωτικής φωσφορυλίωσης. Κάποιο ποσοστό του πυροσταφυλικού μετατρέπεται επίσης σε γλυκόζη από το ήπαρ, το μεγαλύτερο μέρος της οποίας χρησιμοποιείται για να ανακάμψουν τα αποθέματα γλυκογόνου των μυών και του ήπατος τα οποία έχουν εξαντληθεί κατά τη διάρκεια της άσκησης. Η ολοκλήρωση των βιοχημικών αντιδράσεων της επεξεργασίας του πυροσταφυλικού απαιτεί αρκετές ώρες.
Η EPOC επομένως παρέχει το απαιτούμενο Ο2, ώστε να αποκατασταθούν τα αποθέματα φωσφορικής κρεατίνης, να απομακρυνθεί το γαλακτικό και να ανακάμψουν, τουλάχιστον μερικώς, τα αποθέματα γλυκογόνου. Πέρα από την αυξημένη πρόσληψη Ο2, μετά από εξαντλητική άσκηση κατά τη διάρκεια της οποίας παρατηρείται σοβαρή εξάντληση των αποθεμάτων γλυκογόνου (όπως μετά από έναν μαραθώνιο) υπάρχει έντονη ανάγκη αναπλήρωσης των θρεπτικών συστατικών. Σε αυτές τις περιπτώσεις, η ανάκαμψη είναι μακροχρόνια και μπορεί να διαρκέσει μία ημέρα ή περισσότερο, λόγω του ότι η αναπλήρωση των ενεργειακών αποθεμάτων προϋποθέτει την πρόσληψη τροφής. Έτσι, ανάλογα με το είδος και τη διάρκεια της άσκησης, η ανάκαμψη μπορεί να ολοκληρωθεί σε μερικά λεπτά ή μπορεί να απαιτηθεί μία ολόκληρη ημέρα ή περισσότερο.
Μέρος της EPOC δε σχετίζεται ευθέως με την αναπλήρωση των ενεργειακών αποθεμάτων, αλλά οφείλεται σε μια γενικευμένη μεταβολική διαταραχή που έπεται της άσκησης. Για παράδειγμα, η τοπική αύξηση της θερμοκρασίας στους μυς που οφείλεται στην αυξημένη παραγωγή θερμότητας από τη συσταλτική τους δραστηριότητα επιταχύνει όλες τις χημικές αντιδράσεις στον μυϊκό ιστό, συμπεριλαμβανομένων εκείνων που εξαρτώνται από το Ο2. Κατά ανάλογο τρόπο, η θερμοκρασία του σώματος ανεβαίνει κατά αρκετά δέκατα στην κλίμακα Κελσίου στη διάρκεια της άσκησης, γεγονός που επιταχύνει σε ολόκληρο το σώμα τις χημικές αντιδράσεις που καταναλώνουν Ο2. Η αυξημένη ταχύτητα των αντιδράσεων που καταναλώνουν Ο2 μέχρι η θερμοκρασία του σώματος να επανέλθει στα φυσιολογικά (στα πριν από την άσκηση) επίπεδα συμβάλλει στην EPOC.
Επιπλέον, κατά τη διάρκεια της άσκησης αυξάνεται η έκκριση επινεφρίνης, μιας ορμόνης που αυξάνει την κατανάλωση Ο2 από το σώμα. Μέχρι τώρα έχουμε εξετάσει τις συσταλτικές και τις μεταβολικές λειτουργίες των σκελετικών μυϊκών ινών γενικά. Δε χρησιμοποιούν όμως στον ίδιο βαθμό τους παραπάνω μηχανισμούς όλες οι σκελετικές μυϊκές ίνες. Στη συνέχεια θα εξετάσουμε τους τύπους στους οποίους κατατάσσονται οι μυϊκές ίνες, ανάλογα με την ταχύτητα συστολής τους (που εξαρτάται από τον ρυθμό με τον οποίο υδρολύουν την ATP) και τους μηχανισμούς που κυρίως χρησιμοποιούν για να παράγουν ATP.
Οι σκελετικές μυϊκές ίνες κατατάσσονται σε τρεις τύπους με βάση τις διαφορές τους στην υδρόλυση και στη σύνθεση ATP. Με βάση τις βιοχημικές τους ιδιότητες, οι μυϊκές ίνες ταξινομούνται σε τρεις βασικούς τύπους:
- Στις βραδείες οξειδωτικές (τύπου Ι) ίνες
- Στις ταχείες οξειδωτικές (τύπου IIa) ίνες
- Στις ταχείες γλυκολυτικές (τύπου ΙΙx) ίνες
Όπως δηλώνουν τα ονόματά τους, οι δύο κύριες διαφορές μεταξύ των παραπάνω τύπων μυϊκών ινών είναι η ταχύτητα της συστολής τους (βραδεία ή ταχεία) και το είδος των ενζυμικών αντιδράσεων στις οποίες κυρίως βασίζονται για την παραγωγή ATP (οξειδωτική φωσφορυλίωση ή γλυκόλυση).
ΣΥΓΚΡΙΣΗ ΤΑΧΕΩΝ ΚΑΙ ΒΡΑΔΕΩΝ ΜΥΪΚΩΝ ΙΝΩΝ
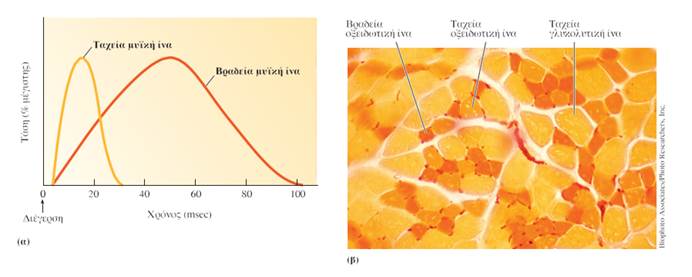
Οι ταχείες μυϊκές ίνες (fast muscle fibers) εμφανίζουν υψηλότερη δραστικότητα ATPάσης της μυοσίνης σε σχέση με τις βραδείες μυϊκές ίνες (slow muscle fibers). Όσο υψηλότερη είναι η δραστικότητα αυτή, τόσο ταχύτερος είναι ο ρυθμός διάσπασης της ATP και επομένως τόσο ταχύτερα διατίθεται η απαιτούμενη ενέργεια για την πραγματοποίηση των επαναλαμβανόμενων κύκλων αλληλεπίδρασης της ακτίνης με τη μυοσίνη. Έτσι συσπώνται ταχύτερα σε σχέση με τις βραδείες μυϊκές ίνες που διασπούν την ATP με βραδύτερο ρυθμό. Κατά μέσο όρο, ο χρόνος για την ανάπτυξη μέγιστης δύναμης συστολής στις ταχείες μυϊκές ίνες επιτυγχάνεται σε 15-40 msec, ενώ στις βραδείες μυϊκές ίνες επιτυγχάνεται σε 50-100 msec.
Δύο λοιπόν παράγοντες καθορίζουν την ταχύτητα συστολής των μυών: το φορτίο που εφαρμόζεται σε αυτούς (σχέση φορτίου-ταχύτητας) και τα επίπεδα δραστικότητας της ATPάσης της μυοσίνης των συστελλόμενων ινών.
ΣΥΓΚΡΙΣΗ ΟΞΕΙΔΩΤΙΚΩΝ ΚΑΙ ΓΛΥΚΟΛΥΤΙΚΩΝ ΜΥΪΚΩΝ ΙΝΩΝ
Οι μυϊκές ίνες διαφέρουν επίσης ως προς την ικανότητά τους να συνθέτουν ATP. Οι μυϊκές ίνες με τη μεγαλύτερη ικανότητα σύνθεσης ATP είναι περισσότερο ανθεκτικές στην κόπωση. Ορισμένες μυϊκές ίνες είναι καλύτερα «εξοπλισμένες» μεταβολικά για την πραγματοποίηση οξειδωτικής φωσφορυλίωσης, ενώ σε άλλες η σύνθεση της ATP πραγματοποιείται κυρίως αναερόβια μέσω της γλυκόλυσης. Τα ενεργειακά αποθέματα των πρώτων διαρκούν περισσότερο, επειδή η οξειδωτική φωσφορυλίωση παράγει σημαντικά περισσότερα μόρια ATP ανά μόριο θρεπτικού συστατικού που διασπάται. Επιπλέον, η παραγωγή ATP μέσω της οξειδωτικής φωσφορυλίωσης δεν οδηγεί σε συσσώρευση γαλακτικού οξέος. Κατά συνέπεια οι αερόβιες μυϊκές ίνες είναι πιο ανθεκτικές στην κόπωση σε σχέση με τις γλυκολυτικές ίνες.
Ορισμένα άλλα χαρακτηριστικά των τριών παραπάνω τύπων μυϊκών ινών που τους διαφοροποιούν μεταξύ τους συνοψίζονται στον ακόλουθο πίνακα.
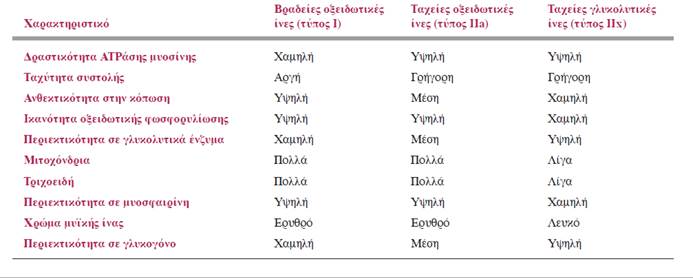
Όπως είναι αναμενόμενο, οι οξειδωτικές μυϊκές ίνες (oxidative muscle fibers), τόσο οι ταχείες όσο και οι βραδείες, περιέχουν σε μεγάλη αφθονία τα οργανίδια στα οποία πραγματοποιείται η οξειδωτική φωσφορυλίωση: τα μιτοχόνδρια.
Επειδή η οξειδωτική φωσφορυλίωση χρειάζεται Ο2, οι οξειδωτικές μυϊκές ίνες αιματώνονται από ένα εκτεταμένο δίκτυο τριχοειδών. Οι οξειδωτικές ίνες περιέχουν επίσης υψηλά ποσά μυοσφαιρίνης. Η μυοσφαιρίνη αυξάνει τη μεταφορά Ο2 σε αυτές και τους προσδίδει κόκκινο χρώμα, όπως η οξυγονωμένη αιμοσφαιρίνη προσδίδει κόκκινο χρώμα στο αρτηριακό αίμα. Οι παραπάνω ίνες ονομάζονται επίσης ερυθρές ίνες (red fibers) λόγω του χρώματός τους.
Αντίθετα, οι ταχείες ίνες που εξαρτώνται από τη γλυκόλυση για την παραγωγή ATP περιέχουν σχετικά λίγα μιτοχόνδρια, ενώ είναι πλούσιες σε γλυκολυτικά ένζυμα. Επίσης, προκειμένου να εξασφαλίσουν τις μεγάλες ποσότητες γλυκόζης που απαιτούνται για τη γλυκόλυση, αυτές οι ίνες περιέχουν μεγάλα αποθέματα γλυκογόνου. Το δίκτυο των τριχοειδών που αιματώνει τις γλυκολυτικές μυϊκές ίνες (glycolytic muscle fibers), όπως αναμένεται, είναι λιγότερο πυκνό, καθώς χρειάζονται σχετικά λιγότερο Ο2 για να παράγουν ενέργεια. Οι γλυκολυτικές ίνες περιέχουν επίσης σχετικά μικρή ποσότητα μυοσφαιρίνης, με αποτέλεσμα να έχουν λευκωπό ή ωχρό χρώμα και γι’ αυτό μερικές φορές ονομάζονται λευκές ίνες (white fibers).
Ο ΤΥΠΟΣ ΤΩΝ ΜΥΪΚΩΝ ΙΝΩΝ ΚΑΘΟΡΙΖΕΤΑΙ ΑΠΟ ΓΕΝΕΤΙΚΟΥΣ ΠΑΡΑΓΟΝΤΕΣ
Στους ανθρώπους, οι περισσότεροι μύες περιέχουν μυϊκές ίνες όλων των τύπων. Το ποσοστό των διαφόρων τύπων ινών σε κάθε μυ εξαρτάται πρωτίστως από τη λειτουργία στην οποία αυτός εξειδικεύεται. Έτσι, οι μύες που χρειάζεται να πραγματοποιούν κυρίως χαμηλής έντασης παρατεταμένες συσπάσεις χωρίς να εμφανίζουν εύκολα κόπωση (όπως οι μύες της ράχης και των ποδιών που στηρίζουν το βάρος του σώματος) έχουν υψηλό ποσοστό βραδέων οξειδωτικών ινών. Οι ταχείες γλυκολυτικές ίνες επικρατούν στους μυς των χεριών, οι οποίοι χρειάζεται να εκτελούν ταχείες και δυναμικές κινήσεις, για παράδειγμα κατά την ανύψωση βαρέων αντικειμένων.
Το ποσοστό των διαφόρων τύπων των μυϊκών ινών δε διαφέρει μόνο μεταξύ των μυών σε ένα άτομο, αλλά και μεταξύ των ατόμων. Αθλητές που είναι γενετικά προικισμένοι με υψηλότερο ποσοστό ταχέων γλυκολυτικών ινών έχουν πλεονέκτημα στους αγώνες ταχύτητας, ενώ αυτοί που έχουν υψηλότερο ποσοστό βραδέων οξειδωτικών μυϊκών ινών είναι πιο πιθανό να υπερέχουν σε αγώνες αντοχής όπως ο μαραθώνιος. Φυσικά, η επιτυχία σε κάθε άθλημα δεν εξαρτάται μόνο από τις γενετικές καταβολές, αλλά και από πολλούς άλλους παράγοντες, για παράδειγμα από τον βαθμό και τον τύπο της προπόνησης.
Πράγματι, οι μηχανικές και οι μεταβολικές ιδιότητες των μυϊκών ινών μπορούν να αλλάξουν σε μεγάλο βαθμό ανταποκρινόμενες στις απαιτήσεις που τους θέτουμε. Οι μυϊκές ίνες προσαρμόζονται σε σημαντικό βαθμό στις λειτουργικές απαιτήσεις που τίθενται σε αυτές. Οι διαφορετικοί τύποι άσκησης προκαλούν διαφορετικά πρότυπα νευρικής διέγερσης στους μυς που εμπλέκονται. Ανάλογα με αυτά τα πρότυπα, λαμβάνουν χώρα μακροχρόνιες προσαρμοστικές αλλαγές στις μυϊκές ίνες, που τις καθιστούν ικανές να ανταποκρίνονται πιο αποτελεσματικά στις λειτουργικές απαιτήσεις που επιβάλλονται στον μυ. Οι αλλαγές που επάγονται στις μυϊκές ίνες αφορούν (1) την ικανότητά τους να πραγματοποιούν αερόβιο μεταβολισμό και (2) τη διάμετρό τους.
ΒΕΛΤΙΩΣΗ ΤΗΣ ΟΞΕΙΔΩΤΙΚΗΣ ΙΚΑΝΟΤΗΤΑΣ
Η τακτική αερόβια άσκηση αντοχής, όπως η κολύμβηση και το τζόκινγκ, προάγει μεταβολικές αλλαγές στις αερόβιες ίνες, καθώς αυτές είναι κυρίως οι ίνες που στρατολογούνται κατά τη διάρκειά της. Για παράδειγμα, αυξάνονται ο αριθμός των τριχοειδών που τους παρέχουν αίμα και ο αριθμός των μιτοχονδρίων τους. Έτσι οι μύες προσαρμόζονται ώστε να χρησιμοποιούν το Ο2 πιο αποτελεσματικά και επομένως καθίστανται ικανότεροι να υποστηρίξουν παρατεταμένη δραστηριότητα χωρίς να εμφανίσουν κόπωση. Σε αυτή την περίπτωση όμως δεν παρατηρείται αλλαγή στο μέγεθος των μυών.
ΜΥΪΚΗ ΥΠΕΡΤΡΟΦΙΑ
Το μέγεθος των μυών μπορεί να αυξηθεί με τακτική, μικρής διάρκειας και υψηλής έντασης αναερόβια άσκηση αντίστασης (resistance training), όπως άρση βαρών. Η αύξηση του μεγέθους των μυών, δηλαδή η υπερτροφία (hypertrophy) τους, σε αυτή την περίπτωση προέρχεται κατά κύριο λόγο από την αύξηση της διαμέτρου των ταχέων γλυκολυτικών ινών, οι οποίες επιστρατεύονται κατά τη διάρκεια έντονων συσπάσεων με μεγάλη δύναμη συστολής. Η αύξηση της διαμέτρου των ταχέων γλυκολυτικών ινών οφείλεται κυρίως στην αυξημένη σύνθεση νηματίων ακτίνης και μυοσίνης, η οποία αποσκοπεί στην αύξηση της δυνατότητας πραγματοποίησης δυναμικών έλξεων και κατά συνέπεια στην αύξηση της συσταλτικής δύναμης του μυός.
Το μηχανικό στρες που ασκεί η άσκηση με βάρη σε μια μυϊκή ίνα ενεργοποιεί πρωτεΐνες μετάδοσης σήματος, οι οποίες ακολούθως ενεργοποιούν τα γονίδια εκείνα που κωδικοποιούν τις πρωτεΐνες που μετέχουν στη συστολή του μυός. Η έντονη άσκηση με βάρη μπορεί να διπλασιάσει ή να τριπλασιάσει τον όγκο ενός μυός. Οι υπερτροφικοί μύες είναι καλύτερα προσαρμοσμένοι στις δραστηριότητες που απαιτούν έντονη σύσπαση για μικρό χρονικό διάστημα, όμως η αντοχή τους δεν παρουσιάζει βελτίωση.
ΕΠΙΔΡΑΣΗ ΤΗΣ ΤΕΣΤΟΣΤΕΡΟΝΗΣ
Οι μυϊκές ίνες των ανδρών έχουν μεγαλύτερη διάμετρο και κατά συνέπεια οι μύες τους είναι μεγαλύτεροι και ισχυρότεροι σε σχέση με τους μυς των γυναικών, ακόμα και χωρίς άσκηση με βάρη. Το γεγονός αυτό οφείλεται στη δράση της τεστοστερόνης, η οποία είναι μια στεροειδής ορμόνη που εκκρίνεται κατά κύριο λόγο στους άνδρες. Η τεστοστερόνη προάγει τη σύνθεση της ακτίνης και της μυοσίνης.
ΑΛΛΗΛΟΜΕΤΑΤΡΟΠΗ ΤΩΝ ΤΑΧΕΩΝ ΜΥΪΚΩΝ ΙΝΩΝ
Όλες οι μυϊκές ίνες μιας κινητικής μονάδας είναι είτε ταχείες είτε βραδείες. Ωστόσο, οι δύο τύποι ινών ταχείας σύσπασης (οι οξειδωτικές και οι γλυκολυτικές) είναι δυνατόν να μετατρέπονται ο ένας στον άλλο, προσαρμοζόμενοι στο είδος της προπόνησης ενός ατόμου, δηλαδή στις λειτουργικές απαιτήσεις που επανειλημμένα επιβάλλονται σε αυτούς τους τύπους ινών από τα κινητικά τους νεύρα. Οι ταχείες και οι βραδείες ίνες δεν μπορούν ωστόσο να μετατραπούν η μία στην άλλη.
ΟΙ ΜΥΕΣ ΔΙΑΘΕΤΟΥΝ ΜΙΑ ΜΙΚΡΗ ΙΚΑΝΟΤΗΤΑ ΕΠΙΔΙΟΡΘΩΣΗΣ
Όταν ένας μυς υποστεί βλάβη, είναι δυνατή μια περιορισμένη αποκατάσταση, παρά το γεγονός ότι τα μυϊκά κύτταρα δεν μπορούν να διαιρεθούν μιτωτικά ώστε να αντικαταστήσουν τα κύτταρα που χάθηκαν. Κοντά στην επιφάνεια των μυών εντοπίζεται ένας μικρός πληθυσμός μυϊκών βλαστικών κυττάρων τα οποία βρίσκονται σε λανθάνουσα κατάσταση και ονομάζονται δορυφορικά κύτταρα (satellite cells).
Όταν μια μυϊκή ίνα υποστεί βλάβη, ελευθερώνονται τοπικά παράγοντες που ενεργοποιούν τα δορυφορικά κύτταρα, τα οποία διαιρούνται δίνοντας γένεση σε μυοβλάστες, σε αυτόν δηλαδή τον τύπο αδιαφοροποίητων κυττάρων που διαφοροποιούνται σε μυϊκές ίνες κατά τη διάρκεια της εμβρυϊκής ανάπτυξης.
Οι μυοβλάστες συγχωνεύονται ανά ομάδες και σχηματίζουν μεγάλα πολυπύρηνα κύτταρα τα οποία ξεκινούν άμεσα τη σύνθεση και τη συναρμολόγηση των ενδοκυτταρικών συστατικών των ώριμων μυϊκών ινών και τελικά διαφοροποιούνται πλήρως σε ώριμες μυϊκές ίνες.
Σε εκτεταμένους τραυματισμούς των μυών, ο παραπάνω μηχανισμός δεν επαρκεί ώστε να αντικατασταθούν πλήρως όλες οι μυϊκές ίνες που χάθηκαν.
ΝΕΚΡΙΚΗ ΑΚΑΜΨΙΑ
Να σημειωθεί ότι ένα νέο μόριο ATP θα πρέπει να δεσμευθεί από τη μυοσίνη, ώστε να είναι δυνατή η αποσύνδεση της εγκάρσιας γέφυρας από το μόριο της ακτίνης στο τέλος κάθε κύκλου δυναμικής έλξης (παρόλο που η ATP δε διασπάται κατά την αποσύνδεση των δύο πρωτεϊνών). Η σημασία της ATP στην αποδέσμευση της μυοσίνης από την ακτίνη γίνεται εμφανής όταν κανείς παρατηρεί το φαινόμενο της νεκρικής ακαμψίας (rigor mortis). Αυτή η «ακαμψία του θανάτου» είναι ένα γενικευμένο «κλείδωμα» των σκελετικών μυών που αρχίζει 3 έως 4 ώρες μετά τον θάνατο και ολοκληρώνεται περίπου σε 12 ώρες. Μετά τον θάνατο, η συγκέντρωση Ca2+ στο κυτταρόπλασμα αρχίζει να αυξάνεται, πιθανότατα λόγω του ότι η ανενεργή πλέον μεμβράνη των μυϊκών κυττάρων δεν μπορεί να κρατήσει έξω από αυτά το εξωκυτταρικό Ca2+, αλλά και λόγω του ότι Ca2+ διαρρέει από τους πλευρικούς σάκους. Οι αυξημένες συγκεντρώσεις Ca2+ προκαλούν τη μετατόπιση της τροπονίνης και της τροπομυοσίνης, επιτρέποντας τη σύνδεση της ακτίνης με τις εγκάρσιες γέφυρες μυοσίνης, οι οποίες έχουν ήδη «φορτιστεί» με ATP πριν από τον θάνατο. Επειδή τα νεκρά κύτταρα δεν μπορούν να παραγάγουν ATP, η μυοσίνη και η ακτίνη δεν μπορούν να αποσυνδεθούν. Τα παχέα και τα λεπτά νημάτια παραμένουν επομένως συνδεδεμένα στις ακινητοποιημένες εγκάρσιες γέφυρες, γεγονός που προκαλεί την ακαμψία των νεκρών μυών. Κατά τη διάρκεια των επόμενων ημερών, η νεκρική ακαμψία σταδιακά υποχωρεί, καθώς αρχίζει η αποδόμηση των μυϊκών πρωτεϊνών.
Έλεγχος της κινητικής δραστηριότητας
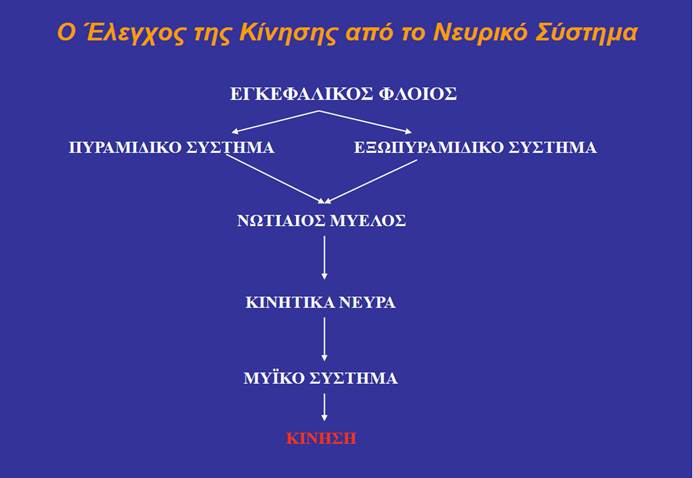
Ο έλεγχος της κινητικής δραστηριότητας διεκπεραιώνεται μέσω τριών κατηγοριών νευρικών σημάτων που μεταβιβάζονται στους κινητικούς νευρώνες των διαφόρων μυών:
(1) σήματα προερχόμενα από νωτιαία αντανακλαστικά τόξα, τα οποία ξεκινούν από προσαγωγούς νευρώνες,
(2) σήματα προερχόμενα από το φλοιονωτιαίο (πυραμιδικό) κινητικό σύστημα, που ξεκινά από τον πρωτοταγή κινητικό φλοιό και ρυθμίζει τις ακριβείς λεπτές κινήσεις των χεριών και
(3) σήματα προερχόμενα από το πολυνευρωνικό (εξωπυραμιδικό) κινητικό σύστημα, το οποίο εκτείνεται από το εγκεφαλικό στέλεχος και ρυθμίζει τη στάση του σώματος για την οποία απαιτούνται ακούσιες κινήσεις μεγάλων ομάδων μυών του κορμού και των άκρων. Τα ερεθίσματα που μεταβιβάζει το εγκεφαλικό στέλεχος επηρεάζονται από την παρεγκεφαλίδα, τα βασικά γάγγλια και τον εγκεφαλικό φλοιό.
Η σωστή ρύθμιση των κινητικών εντολών προϋποθέτει συνεχή πληροφόρηση από προσαγωγά ερεθίσματα και ιδιαίτερα ενημέρωση σχετικά με αλλαγές στο μήκος των μυών (που καταγράφουν οι μυϊκές άτρακτοι) και στην τάση των μυών (που καταγράφουν τα τενόντια όργανα Golgi).
Όταν διατείνεται ολόκληρος o μυς, η διάταση των μυϊκών του ατράκτων ενεργοποιεί το μυοτατικό αντανακλαστικό, το οποίο επάγει τη σύσπαση του μυός. Το αντανακλαστικό αυτό αποτελεί έναν μηχανισμό που ανιχνεύει και περιορίζει την παθητική αλλαγή στο μέγεθος του μυός. Οι κινητικές μονάδες καθορίζουν την κινητική δραστηριότητα, η οποία ποικίλλει από τις μυϊκές συσπάσεις που διατηρούν τη στάση του σώματος και την ισορροπία μέχρι τις στερεοτυπικές κινήσεις (όπως η βάδιση) και τις πολύ επιδέξιες κινήσεις (όπως εκείνες που εκτελούνται στην ενόργανη γυμναστική). Ο έλεγχος όλων των κινήσεων, ανεξάρτητα από το επίπεδο πολυπλοκότητάς τους, καθορίζεται από τα νευρικά ερεθίσματα που λαμβάνουν οι κινητικοί νευρώνες, οι οποίοι στη συνέχεια πυροδοτούν τη συστολή των μυϊκών ινών στις αντίστοιχες κινητικές μονάδες.
H λειτουργία των κινητικών μονάδων ρυθμίζεται από πολλαπλά νευρικά ερεθίσματα.
Τρεις κατηγορίες νευρικών ερεθισμάτων (νευρικών σημάτων) επιδρούν στους κινητικούς νευρώνες, προκειμένου να ρυθμίζουν τη διέγερση από αυτούς των μυϊκών ινών τις οποίες νευρώνουν:
- Ερεθίσματα από προσαγωγούς νευρώνες. Τα ερεθίσματα αυτά συνήθως μεταδίδονται μέσω διάμεσων νευρώνων και λαμβάνουν χώρα στο επίπεδο του νωτιαίου μυελού – αφορούν δηλαδή νωτιαία αντανακλαστικά τόξα.
- Ερεθίσματα από τον πρωτοταγή κινητικό φλοιό. Οι νευρικές ίνες μιας κατηγορίας νευρώνων του πρωτογενούς κινητικού φλοιού, οι οποίοι ονομάζονται πυραμιδικά κύτταρα, συνδέονται απευθείας (χωρίς ενδιάμεσες συνάψεις) με τους κινητικούς νευρώνες (ή με τοπικούς διάμεσους νευρώνες που καταλήγουν στους κινητικούς νευρώνες) του νωτιαίου μυελού. Οι ίνες αυτές αποτελούν το φλοιονωτιαίο (corticospinal) ή πυραμιδικό (pyramidal) κινητικό σύστημα (motor system).
- Ερεθίσματα από το εγκεφαλικό στέλεχος, που αποτελεί τμήμα του λεγόμενου πολυνευρωνικού (multineuronal) ή εξωπυραμιδικού (extrapyramidal) κινητικού συστήματος (motor system). Το εξωπυραμιδικό σύστημα περιλαμβάνει πολλές περιοχές του εγκεφάλου οι οποίες επικοινωνούν μεταξύ τους μέσω ενός εκτεταμένου δικτύου συνάψεων. Το εξωπυραμιδικό σύστημα καταλήγει στο εγκεφαλικό στέλεχος, και ειδικά στον δικτυωτό σχηματισμό, ο οποίος δέχεται ερεθίσματα και επηρεάζεται από τον κινητικό φλοιό, την παρεγκεφαλίδα και τους βασικούς πυρήνες (τα βασικά γάγγλια). Επιπλέον, ο κινητικός φλοιός επικοινωνεί με τον θάλαμο, καθώς και με την προκινητική και τη συμπληρωματική κινητική περιοχή. Όλες οι παραπάνω δομές αποτελούν τμήματα του εξωπυραμιδικού συστήματος.
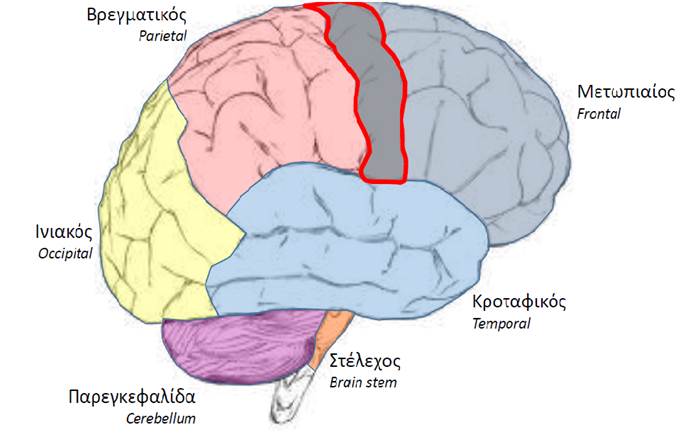
Οι μόνες εγκεφαλικές περιοχές που επηρεάζουν άμεσα τους κινητικούς νευρώνες είναι ο πρωτοταγής κινητικός φλοιός και το εγκεφαλικό στέλεχος. Οι υπόλοιπες περιοχές του εγκεφάλου που εμπλέκονται στον έλεγχο της κινητικής δραστηριότητας επηρεάζουν έμμεσα τους κινητικούς νευρώνες, ρυθμίζοντας τα κινητικά σήματα που διαβιβάζουν ο κινητικός φλοιός και το εγκεφαλικό στέλεχος. Οι διάφορες εγκεφαλικές περιοχές που συμμετέχουν στον έλεγχο της κίνησης αλληλεπιδρούν εκτεταμένα μεταξύ τους.
Τα νωτιαία αντανακλαστικά στα οποία εμπλέκονται προσαγωγοί νευρώνες είναι σημαντικά για τη διατήρηση της στάσης του σώματος και για την εκτέλεση βασικών προστατευτικών κινήσεων, όπως το αντανακλαστικό απόσυρσης.
Το φλοιονωτιαίο σύστημα ελέγχει κατά κύριο λόγο την εκτέλεση λεπτών, διακριτών, εκούσιων κινήσεων των χεριών και των δακτύλων, για παράδειγμα εκείνων που απαιτούνται για να στείλουμε ένα μήνυμα κειμένου από το κινητό μας τηλέφωνο. Η προκινητική και η συμπληρωματική κινητική περιοχή, οι οποίες δέχονται ερεθίσματα από τη νεοπαρεγκεφαλίδα (εγκεφαλοπαρεγκεφαλίδα), προγραμματίζουν τις εκούσιες κινήσεις, μεταβιβάζοντας εντολές στους κατάλληλους κινητικούς νευρώνες μέσω του πρωτογενούς κινητικού φλοιού. Αντίθετα, το εξωπυραμιδικό σύστημα ελέγχει κατά κύριο λόγο τη συνολική στάση του σώματος, για την οποία απαιτούνται ακούσιες κινήσεις μεγάλων ομάδων μυών του κορμού και των άκρων. Το φλοιονωτιαίο και το πολυνευρωνικό σύστημα αλληλεπιδρούν με πολύπλοκο τρόπο και έχουν επικαλυπτόμενες δράσεις. Για να ελέγξουμε εκούσια τους αντίχειρές μας προκειμένου, για παράδειγμα, να στείλουμε ένα μήνυμα κειμένου, υποσυνείδητα τα χέρια μας λαμβάνουν μια συγκεκριμένη στάση που μας επιτρέπει να κρατήσουμε το τηλέφωνο στην κατάλληλη θέση.
Κάποια από τα ερεθίσματα που λαμβάνουν οι κινητικοί νευρώνες είναι διεγερτικά, ενώ άλλα είναι ανασταλτικά. Ο συγχρονισμός των κινήσεων εξαρτάται από την ισορροπία μεταξύ αυτών των δύο τύπων ερεθισμάτων. Οι ακόλουθες μορφές κινητικών δυσλειτουργιών οφείλονται σε ελαττωματικό έλεγχο της κινητικής δραστηριότητας:
-Η απώλεια των ανασταλτικών ερεθισμάτων προς τους κινητικούς νευρώνες μπορεί να οδηγήσει σε σπαστική παράλυση (spastic paralysis), μια κατάσταση που χαρακτηρίζεται από αυξημένο μυϊκό τόνο και ενισχυμένα αντανακλαστικά των άκρων.
-Αντίθετα, η απώλεια των διεγερτικών ερεθισμάτων από τα ανώτερα κέντρα προκαλεί χαλαρή παράλυση (flaccid paralysis). Στην κατάσταση αυτή οι μύες είναι χαλαροί (πλαδαροί) και το άτομο δεν μπορεί να πραγματοποιήσει εκούσιες κινήσεις, αν και τα νωτιαία αντανακλαστικά διατηρούνται.
Βλάβες στον πρωτογενή κινητικό φλοιό στη μία πλευρά του εγκεφάλου, που μπορεί να οφείλονται, για παράδειγμα, σε ένα αγγειακό εγκεφαλικό επεισόδιο, προκαλούν χαλαρή παράλυση στην αντίθετη πλευρά του σώματος. Η κατάσταση κατά την οποία η παράλυση περιορίζεται στη μία μόνο πλευρά του σώματος αναφέρεται ως ημιπληγία (hemiplegia).
Η διακοπή όλων των κατιουσών οδών, όπως συμβαίνει στις περιπτώσεις τραυματικής διατομής του νωτιαίου μυελού, οδηγεί σε χαλαρή παράλυση κάτω από το επίπεδο της διατομής. Η παράλυση και των τεσσάρων άκρων που προκύπτει στις περιπτώσεις που η διατομή εντοπίζεται στα ανώτερα τμήματα του νωτιαίου μυελού ονομάζεται τετραπληγία (quadriplegia), ενώ η παράλυση των κάτω άκρων που προκύπτει στις περιπτώσεις που η διατομή εντοπίζεται στα κατώτερα τμήματα ονομάζεται παραπληγία (paraplegia).
-Η καταστροφή των κινητικών νευρώνων –είτε αφορά τα κυτταρικά σώματα είτε αφορά τις απαγωγές νευρικές ίνες– προκαλεί χαλαρή παράλυση και απώλεια αντανακλαστικών στους μυς που νευρώνουν.
-Βλάβες στην παρεγκεφαλίδα ή στους βασικούς πυρήνες δεν προκαλούν παράλυση αλλά μη συγχρονισμένες αδέξιες κινήσεις και παθολογικά πρότυπα κίνησης. Φυσιολογικά αυτές οι εγκεφαλικές περιοχές εξομαλύνουν τις εκούσιες κινήσεις.
-Βλάβη στις ανώτερες περιοχές του φλοιού (στην προκινητική περιοχή ή στη συμπληρωματική κινητική περιοχή) που εμπλέκονται στον προγραμματισμό της κινητικής δραστηριότητας οδηγούν σε ανικανότητα διαβίβασης των κατάλληλων εντολών και στη μη πραγματοποίηση των επιθυμητών αποτελεσμάτων.
Υποδοχείς των μυών παρέχουν προσαγωγό πληροφορία που είναι απαραίτητη για τον έλεγχο της μυϊκής δραστηριότητας.
Η συγχρονισμένη και στοχευμένη δραστηριότητα των σκελετικών μυών εξαρτάται από προσαγωγά ερεθίσματα από ποικίλες πηγές. Στο απλούστερο επίπεδο, τα προσαγωγά σήματα που υποδεικνύουν ότι το δάκτυλό μας αγγίζει μια καυτή επιφάνεια πυροδοτούν αντανακλαστικά συσταλτική δραστηριότητα στους κατάλληλους μυς των χεριών, ώστε το χέρι μας να απομακρυνθεί από το επιβλαβές ερέθισμα. Σε πιο πολύπλοκο επίπεδο, όταν πρόκειται να πιάσουμε μια μπάλα, τα κινητικά συστήματα στον εγκέφαλό μας θα πρέπει να προγραμματίσουν διαδοχικές κινητικές εντολές οι οποίες θα μετακινήσουν και θα τοποθετήσουν το σώμα μας σωστά για τη σύλληψή της, χρησιμοποιώντας εκτιμήσεις για την κατεύθυνση και την ταχύτητά της σύμφωνα με οπτικά ερεθίσματα. Πολλοί μύες που δρουν ταυτόχρονα ή διαδοχικά σε διαφορετικές αρθρώσεις κινητοποιούνται προκειμένου να αλλάξουν η θέση και η στάση του σώματος ταχύτατα χωρίς να χάνεται η ισορροπία του. Προκειμένου να προγραμματίσει σωστά τη δραστηριότητα των σκελετικών μυών, το ΚΝΣ θα πρέπει να γνωρίζει την αρχική θέση του σώματος. Επιπλέον, θα πρέπει να δέχεται συνεχώς πληροφόρηση για την εξέλιξη της κίνησής του, ώστε να προσαρμόζεται ανάλογα. Ο εγκέφαλος λαμβάνει αυτή την πληροφορία, η οποία είναι γνωστή ως ιδιοδεκτικά ερεθίσματα, από υποδοχείς που βρίσκονται στα μάτια, στις αρθρώσεις, στο αιθουσαίο σύστημα και στο δέρμα, καθώς και από υποδοχείς που βρίσκονται στους ίδιους τους εμπλεκόμενους μυς.
Αν κλείσουμε τα μάτια μας και ενώσουμε τα άκρα του δείκτη του αριστερού και του δείκτη του δεξιού μας χεριού σε οποιοδήποτε σημείο στον χώρο, έχουμε ένα απλό παράδειγμα της λειτουργίας των ιδιοδεκτικών υποδοχέων στους μυς και στις αρθρώσεις μας. Είμαστε σε θέση να κάνουμε την παραπάνω κίνηση χωρίς να βλέπουμε, γιατί ο εγκέφαλός μας πληροφορείται συνεχώς για τη θέση των χεριών και των άλλων τμημάτων του σώματος από προσαγωγά ερεθίσματα με τα οποία τον τροφοδοτούν οι υποδοχείς των μυών και των αρθρώσεων.
Δύο τύποι υποδοχέων των μυών καταγράφουν τις μεταβολές στο μήκος και στην τάση τους: οι μυϊκές άτρακτοι και τα τενόντια όργανα Golgi. Το μήκος των μυών καταγράφεται από τις μυϊκές ατράκτους, ενώ οι μεταβολές στην τάση τους παρακολουθούνται από τα τενόντια όργανα Golgi. Και οι δύο παραπάνω τύποι υποδοχέων ενεργοποιούνται από τη διάταση των μυών, αλλά μεταφέρουν διαφορετικές πληροφορίες.
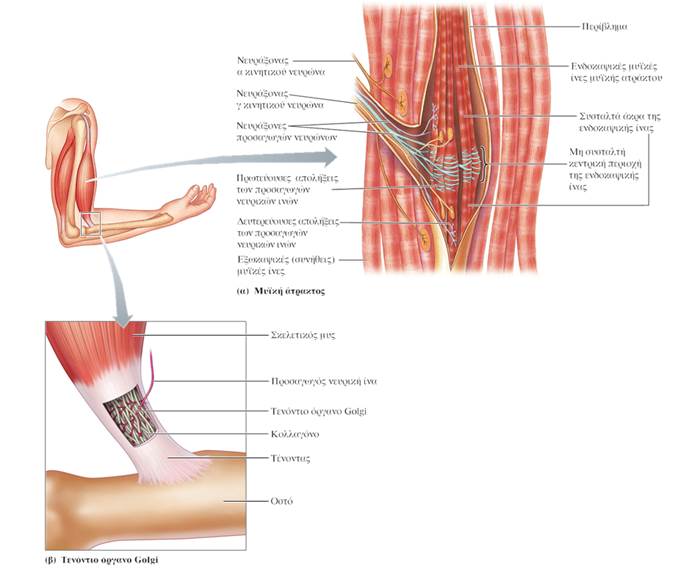
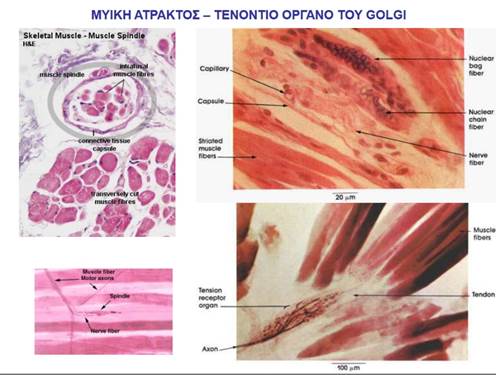
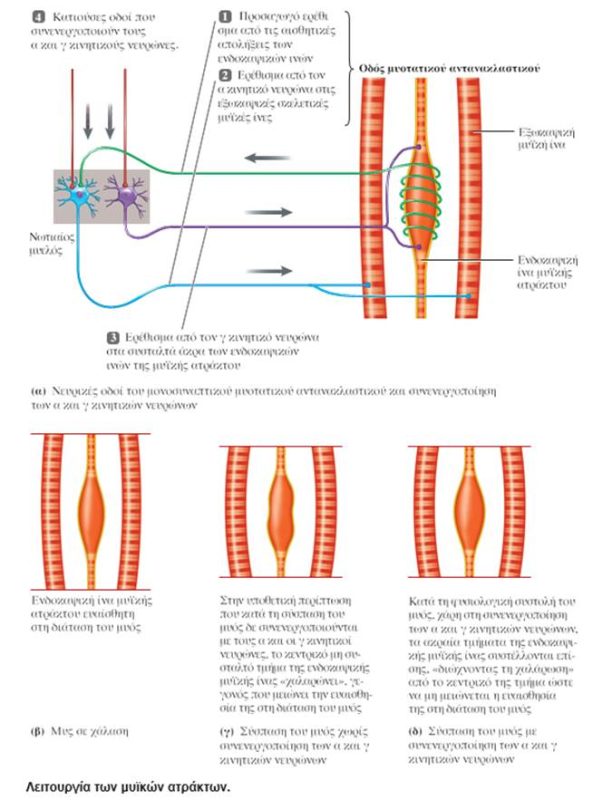
ΔΟΜΗ ΤΩΝ ΜΥΪΚΩΝ ΑΤΡΑΚΤΩΝ
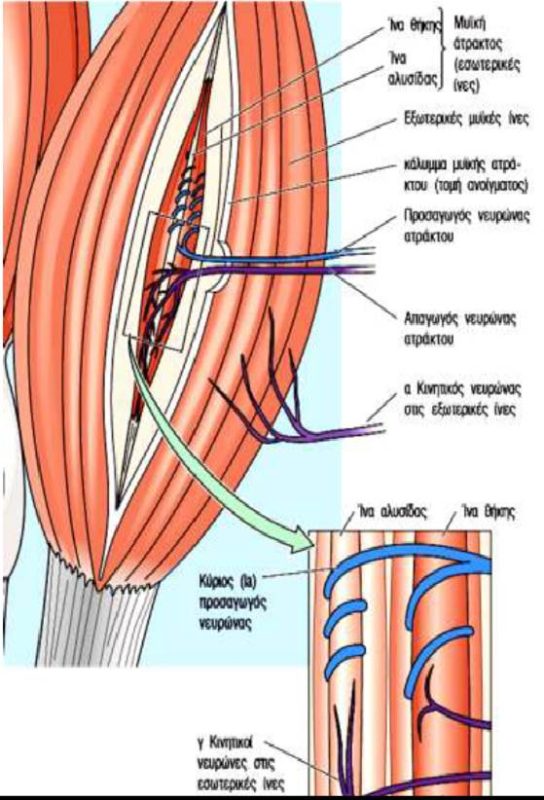
Οι μυϊκές άτρακτοι (muscle spindles) κατανέμονται σε ολόκληρο το σαρκώδες τμήμα των μυών και αποτελούν συναθροίσεις εξειδικευμένων μυϊκών ινών, που είναι γνωστές ως ενδοκαψικές μυϊκές ίνες (intrafusal muscle fibers) και καλύπτονται από ένα περίβλημα συνδετικού ιστού που έχει σχήμα ατράκτου. Οι ενδοκαψικές μυϊκές ίνες διατάσσονται παράλληλα με τις «συνήθεις» μυϊκές ίνες του μυός, τις εξωκαψικές μυϊκές ίνες (extrafusal muscle fibers). Σε αντίθεση με τις εξωκαψικές μυϊκές ίνες, οι οποίες περιέχουν συσταλτά στοιχεία (μυϊκά ινίδια) σε όλο το μήκος τους, οι ενδοκαψικές μυϊκές ίνες περιέχουν μία μη συσταλτή κεντρική περιοχή, ενώ τα μυϊκά ινίδια περιορίζονται στα άκρα τους.
Κάθε μυϊκή άτρακτος διαθέτει ανεξάρτητη νεύρωση από προσαγωγές και απαγωγές νευρικές ίνες. Ο απαγωγός νευρώνας που παρέχει νεύρωση στις ενδοκαψικές μυϊκές ίνες της μυϊκής ατράκτου είναι γνωστός ως γ κινητικός νευρώνας (gamma motor neuron), ενώ οι κινητικοί νευρώνες που παρέχουν νεύρωση στις εξωκαψικές ίνες είναι γνωστοί ως α κινητικοί νευρώνες (alpha motor neurons). Δύο τύποι προσαγωγών αισθητικών απολήξεων καταλήγουν στις ενδοκαψικές ίνες και λειτουργούν ως αισθητήρες των μυϊκών ατράκτων. Και οι δύο αυτοί τύποι διεγείρονται από τη διάταση των μυών. Οι πρωτεύουσες απολήξεις (primary endings) περιελίσσονται γύρω από το κεντρικό τμήμα των ενδοκαψικών ινών και ανιχνεύουν μεταβολές του μήκους των ινών κατά τη διάτασή τους, καθώς και την ταχύτητα της διάτασης. Οι δευτερεύουσες απολήξεις (secondary endings) εντοπίζονται στα ακραία τμήματα πολλών ενδοκαψικών ινών και είναι ευαίσθητες μόνο σε αλλαγές του μήκους των μυών. Οι μυϊκές άτρακτοι διαδραματίζουν βασικό ρόλο στο μυοτατικό αντανακλαστικό.
ΜΥΟΤΑΤΙΚΟ ΑΝΤΑΝΑΚΛΑΣΤΙΚΟ
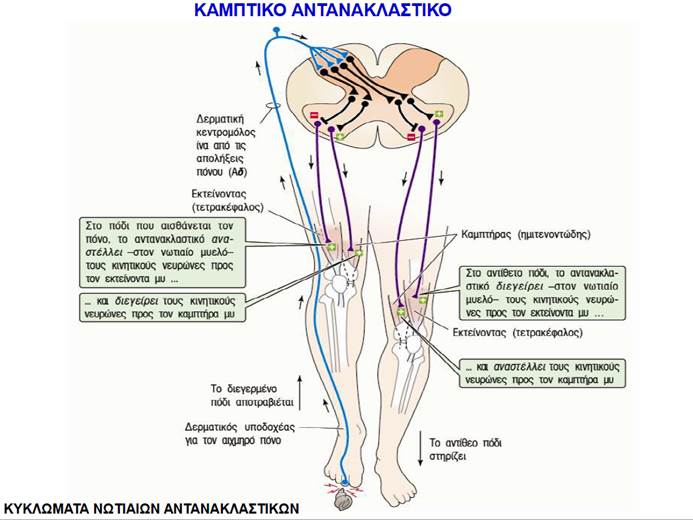
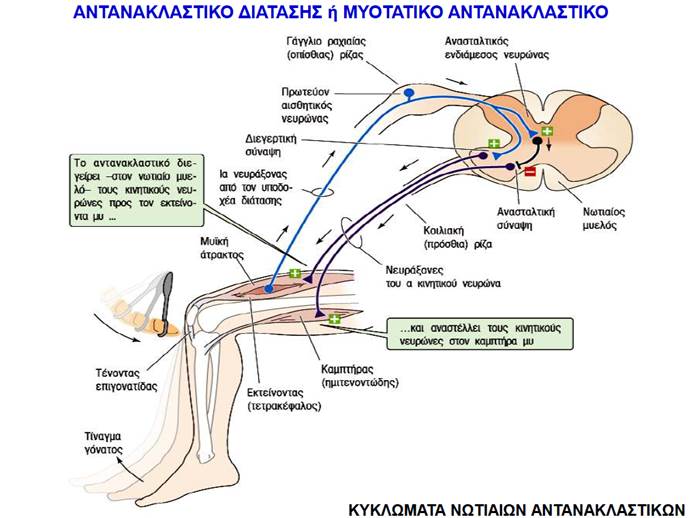
Όταν ένας ολόκληρος μυς διατείνεται παθητικά, οι ενδοκαψικές μυϊκές ίνες των μυϊκών του ατράκτων διατείνονται επίσης, με αποτέλεσμα την αύξηση του ρυθμού πυροδότησης στις προσαγωγές νευρικές ίνες που τις νευρώνουν. Ο προσαγωγός νευρώνας συνάπτεται άμεσα με τον α κινητικό νευρώνα που νευρώνει τις εξωκαψικές ίνες στον ίδιο μυ, γεγονός που προκαλεί τη συστολή του μυός. Αυτό το μονοσυναπτικό μυοτατικό αντανακλαστικό (stretch reflex) αποτελεί έναν τοπικό μηχανισμό αρνητικής ανάδρασης που ανιχνεύει και εμποδίζει τις μεταβολές του μήκους των μυών όταν εφαρμόζεται σε αυτούς πρόσθετο φορτίο.
Το κλασικό παράδειγμα μυοτατικού αντανακλαστικού είναι το αντανακλαστικό της επιγονατίδας (patellar reflex) ή αντανακλαστικό του γόνατος (knee reflex). Ο μυς που εκτείνει την κνήμη είναι ο τετρακέφαλος μηριαίος, ο οποίος σχηματίζει το πρόσθιο τμήμα του μηρού και καταφύεται κάτω από το γόνατο στην κνήμη μέσω του τένοντα της επιγονατίδας (patellar tendon ). Ελαφρύ χτύπημα του τένοντα αυτού με ελαστικό σφυρί διατείνει παθητικά τον τετρακέφαλο μυ, ενεργοποιώντας τις μυϊκές του ατράκτους. Έτσι ενεργοποιείται το μυοτατικό αντανακλαστικό της επιγονατίδας, το οποίο προκαλεί σύσπαση του τετρακεφάλου, με αποτέλεσμα την απότομη έκταση της κνήμης και την ανύψωση του τμήματος του ποδιού που βρίσκεται κάτω από την άρθρωση του γόνατος.
Ο έλεγχος του αντανακλαστικού της επιγονατίδας αποτελεί μέρος της βασικής νευρολογικής εξέτασης. Όταν λειτουργεί φυσιολογικά, σημαίνει πως μια σειρά βασικών νευρικών και μυϊκών στοιχείων –οι μυϊκές άτρακτοι, τα προσαγωγά ερεθίσματα, ο κινητικός νευρώνας, οι απαγωγές νευρικές ώσεις, η νευρομυϊκή σύναψη– λειτουργούν φυσιολογικά. Επίσης, υποδηλώνει ότι τα διεγερτικά και τα ανασταλτικά σήματα με τα οποία τροφοδοτούν τους κινητικούς νευρώνες τα ανώτερα εγκεφαλικά κέντρα βρίσκονται στη σωστή ισορροπία. Τέτοια αντανακλαστικά μπορεί να απουσιάζουν ή να είναι κατεσταλμένα σε περιπτώσεις απώλειας των διεγερτικών ερεθισμάτων από τα ανώτερα εγκεφαλικά κέντρα ή μπορεί να είναι σημαντικά αυξημένα σε περιπτώσεις απώλειας των ανασταλτικών σημάτων προς τους κινητικούς νευρώνες από τα ανώτερα εγκεφαλικά κέντρα.
Ο κύριος ρόλος του αντανακλαστικού της επιγονατίδας είναι η αντίδραση σε φορτία που διατείνουν τους εκτείνοντες μυς των κάτω άκρων. Οποτεδήποτε το γόνατό μας λυγίζει ενώ στεκόμαστε, περπατάμε, τρέχουμε ή πηδάμε, ο τετρακέφαλος μυς διατείνεται. Η διάτασή του, χάρη στο μυοτατικό αντανακλαστικό, προκαλεί ενίσχυση της σύσπασής του και αυτό διατηρεί το πόδι μας ευθειασμένο, ώστε να μπορούμε να παραμένουμε σε όρθια θέση. Ένα παρόμοιο μυοτατικό αντανακλαστικό στον δικέφαλο βραχιόνιο μυ κινητοποιείται όταν πιάνουμε ένα αντικείμενο με το χέρι μας ή όταν γεμίζουμε με νερό ένα ποτήρι που κρατάμε. Στις περιπτώσεις αυτές, η διάταση των μυϊκών ατράκτων του δικεφάλου ενεργοποιεί ένα μυοτατικό αντανακλαστικό σε αυτόν τον μυ που μας βοηθά να κρατήσουμε το αντικείμενο ή να συνεχίσουμε να κρατάμε το ποτήρι καθώς αυτό γίνεται βαρύτερο ενώ το γεμίζουμε με νερό.
ΣΥΝΕΝΕΡΓΟΠΟΙΗΣΗ ΤΩΝ α ΚΑΙ γ ΚΙΝΗΤΙΚΩΝ ΝΕΥΡΩΝΩΝ
Οι γ κινητικοί νευρώνες επάγουν τη συστολή των ακραίων τμημάτων των ενδοκαψικών ινών. Η συστολή αυτή είναι πολύ ασθενής για να έχει κάποια επίδραση στη συνολική τάση που αναπτύσσεται στον μυ, αλλά έχει σημαντική τοπική επίδραση στη μυϊκή άτρακτο. Αν δεν υπήρχαν αντιρροπιστικοί μηχανισμοί, η βράχυνση όλου του μυός από τη διέγερση των α κινητικών νευρώνων που νευρώνουν τις εξωκαψικές ίνες θα «ξεχείλωνε» τις ίνες των μυϊκών ατράκτων, με αποτέλεσμα να καθίσταντο λιγότερο ευαίσθητες στη διάταση και επομένως όχι τόσο αποτελεσματικές ως αισθητήρες του μήκους των μυών. Η συνενεργοποίηση (coactivation) των γ κινητικών νευρώνων μαζί με τους α κινητικούς νευρώνες κατά τη διάρκεια αντανακλαστικών και εκούσιων συσπάσεων προσαρμόζει το μήκος των μυϊκών ατράκτων στη βράχυνση του μυός. Έτσι οι μυϊκές άτρακτοι διατηρούν την ίδια υψηλή ευαισθησία στη διάταση του μυός σε μεγάλο εύρος μηκών αυτού. Η διέγερση των γ κινητικών νευρώνων πυροδοτεί την ταυτόχρονη συστολή των δύο άκρων των ενδοκαψικών ινών, προκαλώντας τη διάταση των κεντρικών, μη συσταλτών τμημάτων τους. Η συστολή των δύο άκρων των ενδοκαψικών ινών «παίρνει τα μπόσικα» που δημιουργεί στις μυϊκές ατράκτους η βράχυνση όλου του μυός. Ο βαθμός διέγερσης των α κινητικών νευρώνων καθορίζεται από την επιζητούμενη δύναμη της μυϊκής συστολής, ενώ ο βαθμός της ταυτόχρονης διέγερσης των γ κινητικών νευρώνων στον ίδιο μυ καθορίζεται από την προβλεπόμενη βράχυνση του μυός. Όταν η βράχυνση όλου του μυός είναι μικρότερη από την αναμενόμενη (για παράδειγμα, το φορτίο είναι μεγαλύτερο από το αναμενόμενο), οι μυϊκές άτρακτοι στέλνουν στους α κινητικούς νευρώνες κατάλληλα σήματα, ώστε να αυξήσουν τον ρυθμό πυροδότησής τους και έτσι να ανταποκριθούν στο αυξημένο φορτίο.
ΤΕΝΟΝΤΙΑ ΟΡΓΑΝΑ GOLGI
Σε αντίθεση με τις μυϊκές ατράκτους, οι οποίες εντοπίζονται στο σαρκώδες κεντρικό τμήμα των μυών (στη μυϊκή γαστέρα), τα τενόντια όργανα Golgi (Golgi tendon organs) εντοπίζονται στους τένοντες των μυών και αντιδρούν όχι σε αλλαγές του μήκους τους, αλλά σε αλλαγές στην τάση που αναπτύσσουν. Επειδή η τάση που αναπτύσσεται συνολικά από έναν μυ κατά τη διάρκεια της συστολής του καθορίζεται από ποικίλους παράγοντες (για παράδειγμα, τη συχνότητα διέγερσής του και το μήκος του μυός κατά την έναρξη της συστολής), είναι απαραίτητο τα κέντρα κινητικού ελέγχου να συλλέγουν πληροφορίες για τη δύναμη που πραγματικά αναπτύσσεται κατά τη διάρκεια μιας συστολής, ώστε να γίνονται οι απαραίτητες προσαρμογές.
Κάθε τενόντιο όργανο Golgi αποτελείται από τις απολήξεις μιας προσαγωγού νευρικής ίνας οι οποίες εισχωρούν στις δεσμίδες ινιδίων κολλαγόνου του συνδετικού ιστού που συνιστά τον τένοντα και περιπλέκονται με τα ινίδια. Όταν οι εξωκαψικές μυϊκές ίνες συσπώνται, δημιουργείται τάση που έλκει τον τένοντα και διατείνει τις δεσμίδες κολλαγόνου του, με αποτέλεσμα την αύξηση της τάσης που ασκείται στο οστό στο οποίο ο τένοντας προσφύεται. Κατά τη διαδικασία αυτή, οι προσαγωγές νευρικές απολήξεις των οργάνων Golgi, οι οποίες περιπλέκονται με τα ινίδια κολλαγόνου, διατείνονται επίσης, γεγονός που επάγει την παραγωγή νευρικών ώσεων από τις προσαγωγές ίνες, με τη συχνότητα πυροδότησής τους να είναι ευθέως ανάλογη με την τάση που αναπτύχθηκε. Η προσαγωγός αυτή πληροφορία μεταβιβάζεται στον εγκέφαλο για επεξεργασία. Σε μεγάλο βαθμό η παραπάνω πληροφορία χρησιμοποιείται ασυνείδητα προκειμένου να εξασφαλιστεί η ομαλότητα των εκτελούμενων κινήσεων αν και, σε αντίθεση με την προσαγωγό πληροφορία από τις μυϊκές ατράκτους, η προσαγωγός πληροφορία από τα τενόντια όργανα Golgi φτάνει το επίπεδο της συνειδητής επίγνωσης. Έτσι αντιλαμβανόμαστε τις μεταβολές στην τάση ενός μυός, ενώ δεν αντιλαμβανόμαστε τις μεταβολές στο μήκος του.
Οι επιστήμονες παλαιότερα πίστευαν ότι τα τενόντια όργανα Golgi ενεργοποιούν ένα προστατευτικό νωτιαίο αντανακλαστικό, το οποίο αναστέλλει την περαιτέρω σύσπαση και επιφέρει την απότομη χαλάρωση των μυών όταν η τάση σε αυτούς αυξάνεται υπερβολικά, ώστε να αποτρέπεται η πρόκληση βλαβών στους μυς και στους τένοντες. Ωστόσο, σήμερα θεωρείται ότι τα τενόντια όργανα Golgi λειτουργούν αποκλειστικά ως αισθητήρες και δεν ενεργοποιούν κανένα αντανακλαστικό. Προφανώς υπάρχουν άλλοι, άγνωστοι προς το παρόν μηχανισμοί οι οποίοι εμπλέκονται στην αναστολή της περαιτέρω σύσπασης των μυών, ώστε να μην αναπτύσσεται υπερβολική τάση που θα μπορούσε να επιφέρει βλάβες.